Vermehrung und Wachstum
V 1.49A
R. Bauer

Wien, Feb. 2015
R. Bauer
Haftungsausschluss: Der Autor übernimmt keinerlei Gewähr für Qualität der bereitgestellten Informationen in diesem Werk.
Haftungsansprüche gegen den Autor, welche sich auf Schäden materieller oder ideeller Art beziehen, die durch die Nutzung oder Nichtnutzung
der dargebotenen Informationen bzw. durch die Nutzung fehlerhafter und unvollständiger, direkte oder referenzierter Informationen verursacht
wurden, sind grundsätzlich ausgeschlossen.
Im Jahr 1768 beschreibt Philip Miller die Keschde im Gardeners Dictionary als Castanea sativa. Sie erhält sein botanisches Kürzel mill (Castanea sativa mill). Der griechische Philosoph und Naturforscher Theophrastos von Eresos nennt die Frucht Euböische Nuss (κάρυον ευβοϊκή).
Die Kultivierung des Baumes zur Nahrungsmittelproduktion dürfte im 9. bis 7. Jahrhundert vor Chr. im Bereich zwischen Kaspischem und Schwarzem Meer erfolgt sein. Von dort verbreitete sich die kultivierte Edelkastanie über Kleinasien, Griechenland und auf den Balkan in Richtung Westen aus. Möglicherweise wurden Edelkastanien auch von den Kelten in Mitteleuropa eingeführt. Jedenfalls verbreiteten die Römer den Weinbau zusammen mit der Edelkastanie im gesamten römischen Reich, wobei der Baum jedoch primär zur Holzgewinnung genutzt wurde. Die Pflanzung der Edelkastanie in Mittel– und Südeuropa, vornehmlich zur Nahrungsmittelerzeugung, erfolgte erst nach den Römern.
Im frühen Mittelalter war die Edelkastanie im südlichen Europa ein wichtiges Nahrungsmittel. Ende des 8. Jahrhunderts oder am Anfang des 9. Jahrhunderts ordnete Karl der Große in seiner Capitulare de villis, die detaillierte Vorschriften über die Verwaltung der Krongüter enthält, auch den Anbau der Castanearios in den kaiserlichen Gütern an.
Die Edelkastanie ist eine langlebige Pflanze, sie erreicht in Europa unter guten Bedingungen ein Alter von 500 – 600 Jahren. Es gibt auch Exemplare, die auf ein weitaus höheres Alter geschätzt werden. Sie kann bis zu 35 m hoch, einen Stammdurchmesser von 2 m und eine Kronenbreite von 30 m aufweisen.
Es gibt in Europa mehrere hundert Sorten, wobei auch Einkreuzungen mit den japanischen und chinesischen Arten (Castanea crenata und Castanea mollissima) gemacht wurden, um vor allem die Resistenz gegen Krankheiten zu erhöhen.
Nicht zu verwechseln ist die Edelkastanie mit der Rosskastanie (Aesculus Hippocastanum), die mit der Edelkastanie nicht verwandt ist. Diese wurde erst 1576 von Baron Ungnad von Konstantinopel nach Wien gesandt, von Carolus Clusius angepflanzt und dann über ganz Europa verbreitet.
Die Edelkastanie Castanea sativa gehört zu den Buchengewächsen (Fagaceae) genauso wie die Eiche (Quercus).
Das enthaltene Wissen erhebt keinen Anspruch auf Vollständigkeit und Richtigkeit im fachlichen Sinn.
Im folgenden Text soll auf die Standortansprüche (Wasser, Temperatur, Licht, Boden), Symbiosen, Schädlinge und auf die Vermehrung mit einfachen Mitteln eingegangen werden. Der Inhalt ist mehrheitlich aus Informationen aus dem Web, aus Gesprächen mit Sachkundigen und auch aus praktischer Erfahrung entstanden.
Es wird im Text auf eine Methode eingegangen, welche auch praktisch erprobt und etwas optimiert wurde. Innerhalb einer Wachstumsperiode lässt sich ein veredelter Baum kultivieren, der bis zu einem Meter Höhe erreicht, einen gut entwickelten Wurzelstock und feste Herbstknospen aufweist.
Um diese Methode erfolgreich anzuwenden, braucht es kein Gewächshaus, Kältekammer oder Luftbefeuchtungsanlage. Als Einstieg reichen Materialien und Werkzeuge aus dem Gartenbedarf bzw. Baumarkt, Aststückchen mit Knospen eines kultivierten Baumes und ein paar Edelkastanien 1.
Und natürlich Freude daran sich Wissen anzueignen und auch etwas Geduld.
Vorschläge, Verbesserungen, Berichtigungen und Erweiterungen bitte an maroni@upwind·at senden. Das Dokument ist auf http://maroni.upwind·at zu finden.
Die Form der ausgewachsenen Blätter ist länglich, etwa 10–25 cm lang und bis zu 8 cm breit (Abbildung 2). Der Blattrand ist gezahnt bis gekerbt. Die voll entwickelten Blätter sind etwas ledrig, die Oberseite des Blattes ist glänzend, tiefgrün und glatt, die Unterseite ist heller, hier treten die Adernpaare klar hervor. Die Herbstfärbung ist gelbbraun, direkt vor dem Abfallen braun. Die Blätter von jungen, noch nicht fruchttragenden Bäumen, bleiben hellgrün und weich – laut [EE93] ein Altersdimporphismus.
Das bedeutet aber auch, dass veredelte Jungbäume dunklere Blätter entwickeln, als sie bei unveredelten Jungbäumen zu finden sind (Abbildung 3)!
In Abbildung 4 werden Rosskastanien mit Blattoberseite und Unterseite gezeigt. In der Abbildung rechts werden Edelkastanien inklusive Igeln und Blättern gezeigt. Im Gegensatz zur Rosskastanie hat die Edelkastanie auf der Frucht eine kleine behaarte Spitze mit Narbenästen!
Abbildung 4: Gegenüberstellung: Rosskastanie li und Edelkastanie re
Abbildung 5: Gegenüberstellung Schnitte: Rosskastanie li und Edelkastanie re
Die Borke (Abbildung 6) ist bei jungen Stämmen noch glatt, die Farbe silbrig-grau. Im Alter wird sie graubraun und reißt auf. Es bildet sich eine dicke Borke mit breiten Streifen.
Abbildung 7: Blüten li und reife Früchte re
Abbildung 8: Edelkastanie als Waldbaum
Ich stehe in einer Gartenabteilung und der Maronibaum vor mir gefällt mir so gut und ist außerdem ein Schnäppchen – ich möchte ihn unbedingt haben!
1. Habe ich genug Platz oder habe ich in nächster Zeit genug Platz um Bäume zu pflanzen?
2. Ist der Pflanzboden für den Baum leicht sauer (pH-Wert um die 6 herum)?
3. Hab ich ein Temperaturjahresmittel von mehr als 10 ∘C?
4. Ist mir bewusst, dass ich zwei mindestens zwei Bäume benötige, wenn es in der Nachbarschaft keinen zweiten Baum gibt, damit die Igeln auch
gefüllte Früchte tragen (Ausnahme: zwei verschiedene Sorten am gleichen Baum)?
5. Ist der Baum veredelt oder anders vegetativ vermehrt?
6. Steht der Baum fest im Topf?
7. Ist die Veredelungsstelle sauber verwachsen?
8. Sieht der Baum gesund aus (Ausscheidungskriterien: Rinde aufgesprungen, Blätter welk, Äste abgebrochen, Spitze verletzt)?
9. Kommt der Baum aus dem eigenen Land?
10. Ist auf dem Schild die genaue Sorte angegeben (z.B. österreichische Sorten wie Ecker1, Ecker2, französische wie Bouche de Bétizac, Bournette, Dorée de Lyon, ...) ?
Oder vielleicht doch zunächst das Dokument einmal durchlesen und dann eine Baumschule aufsuchen,
die Edelkastanienbäume selbst züchtet.
Ich habe einen Baum im Ausland gesehen – er wurde mir günstig angeboten, soll ich ihn mitnehmen? – Eher nicht
Ich habe am Markt Maroni gesehen, die schon austreiben – können daraus Bäume werden? Wenn die austretenden Keimwurzeln noch nicht dunkel sind, dann Ja. Zuhause lockere Erde in einen Topf geben und je eine Maroni flach, jedenfalls mit der Keimwurzel nach unten gerichtet, in einen Topf legen und mit etwas Erde bedecken und nur mäßig gießen. Mit etwas Licht und bei normaler Wohnraumtemperatur gibt es in spätestens vier Wochen einen kleinen Baum. Bis dahin vielleicht doch noch das ganze Dokument durchlesen.
Die Edelkastanie ist ein Tiefwurzler mit intensivem, weit reichendem Wurzelsystem. Der beste Boden für die Edelkastanie ist tiefgründig (mehr als 70 cm) und bietet eine natürliche Drainage. Daher sind auch Hanglagen gut geeignet. Abdichtender Unterboden mit hohem Tonanteil und verdichteter Boden ist weniger geeignet. Der Unterboden von größeren natürlichen Edelkastanienbeständen ist meist sandiger Lehm oder verwittertes Lavagestein.
Der Boden sollte leicht sauer bei einem pH-Wert von 4,5 – 6,5 sein; wobei der bevorzugte pH–Wert bei 5,5 – 6,0 liegt. Silikatböden (Lavagestein, sandiger Lehm, Sandstein, Granit, Gneis, Schiefer) sind als Untergrund geeignet. Kalkböden sind nicht geeignet – außer auf Kalkböden im Untergrund mit genügend hoher Humusauflage. Der Feinkalkgehalt im Wurzelbereich sollte jedenfalls unter 3 % liegen.
Anmerkungen: Auf kalkhaltigen Böden und auch auf staunassen Böden treten Chlorosen an den Blättern auf. Bei etwas kalkhältigen Böden kann eine regelmäßige Torf- oder Rindenauflage oder auch Ammoniumsulfat als Mineraldünger den Kalk zum Teil binden und damit den pH-Wert senken. Auf Kleinflächen können auch spezielle Eisendünger eine Lösung sein um Chlorosen zu verhindern (siehe auch Abschnitt 17.5.4).
Im Kalkstein ist Calciumcarbonat der Hauptbestandteil des sedimentär gebildeten Calcit und Aragonit und so kann eiszeitliches Geschiebe aus den Kalkalpen, viel gebundenen Kalk enthalten. Daher können auch Ablagerungen von Lehm – im Gegensatz zu Tiefenlehm – hohe Anteile von Kalk enthalten.
Landwirtschaftliche Böden und Grünflächen werden nach Nährstoffgehalt in Klassen [NW12]. Die Tabelle ist in Reinnährstoffklassen für Grünland bzw. mittelschwere Böden [Bau06] gegliedert:
| Gehaltsklasse | Phosphor [mg/kg] | Kalium [mg/kg] | Magnesium [mg/kg] |
| A sehr niedrig | unter 26 | unter 50 | unter 30 |
| B niedrig | 26–46 | 50–87 | 30–55 |
| C anzustreben | 47–68 | 88–170 | 56–105 |
| D hoch | 69–274 | 171–332 | 106–190 |
| E sehr hoch | über 274 | über 332 | über 190 |
Für Massenprozentwerte für Trockensubstanz sind die angegebenen Werte durch 10000 zu dividieren – z.B. für Nährstoffgehalt
A und Masse–Phosphoranteil errechnet 0,005 %. Achtung, früher wurde der Nährstoffgehalt in mg/100 g angegeben!
Der Düngungsbedarf an Phosphor, Kalium und Magnesium muss nicht mit so hoher Genauigkeit erfolgen wie der Stickstoffbedarf. Da P, K und Mg im Boden (Abschnitt 17.2.2) gepuffert wird, führt eine überhöhte Düngung normalerweise nicht zu Schäden an den Pflanzen oder zu einer Beeinträchtigung der Umwelt.
Trockener sandiger Lehm wiegt grob abgeschätzt im Mittel 2,7 kg/l. Trockener gewachsener Boden im Mittel etwa 1,7 kg/l. Um als ganz grober Richtwert die gleiche
Nährstoffdichte pro Volumseinheit (von mg/kg auf mg/l) für Substrate/Böden zu erreichen, werden die Tabellenwerte mit 1,7 multipliziert. Für die
Gehaltsklasse C mit 47–68 mg/kg Phosphor ergeben sich 79,9 mg/l–115,6 mg/l Phosphor. Der Vollständigkeit wird nun in der folgenden Tabelle der
Nährstoffgehalt A – E umgerechnet auf die Phosphat/Oxidformen angegeben:
| Gehaltsklasse | Phosphorpentoxid | Kaliumoxid | Magnesiumoxid |
| (P2O5) [mg/kg] | (K2O) [mg/kg] | (MgO) [mg/kg] | |
| A sehr niedrig | unter 60 | unter 60 | unter 50 |
| B niedrig | 60–105 | 60–105 | 50–91 |
| C anzustreben | 106–156 | 106–205 | 92–174 |
| D hoch | 157–528 | 206–400 | 175–315 |
| E sehr hoch | über 528 | über 400 | über 315 |
Bodenanalysen kann man z.B. bei [Reb15] durchführen lassen. Für ertragreiche Edelkastanienbäume sind Böden mit Nährstoffgehalt der Klasse C anzustreben (siehe auch Abschnitt 4).
Die Edelkastanie liebt warmes Klima. Sechs Monate im Jahr sollten über +10 ∘C liegen, darunter werden die Früchte nicht ausreifen. Die Mindestniederschlagsmenge liegt bei 600 mm pro Jahr. Sie ist eine submontan-mediterrane Baumart.
Natürliche Bestände der Edelkastanie in Niederösterreich gedeihen meist auf Hügelkuppen oder auf flach abfallenden Hänge mit NNW, NNO, NO Ausrichtung. Auf Streuobstwiesen im Burgenland in Richtung NO oder Kuppen in der Ebene. Und auf Plantagen in der Steiermark findet man Edelkastanien auf NO Hängen und eine auf einem SSW Hang mit einer Abdeckung durch einen im Süden vorgelagerten Hochwald.
Die Edelkastanie benötigt als Halbschattenbaumart doch relativ viel Licht. Edelkastanienbäume im Topf sollten möglichst nicht dem Frost ausgesetzt werden oder in die pralle Sonne gestellt werden – besonders dunkle Töpfe erhitzen sehr schnell. Wind-exponierte Lagen sind als Pflanzplatz ungeeignet.
Als groben Richtwert verträgt die tief verwurzelte Edelkastanie (Castanea sativa) Wintertemperaturen ohne Schäden bis zu −20 ∘C. Die Winterhärte ist aber arten– und damit auch sortenabhängig. Es gibt auch Anpassungen der gleichen Art an die jeweiligen lokalen klimatische Bedingungen [GGHS11]. Die Frostempfindlichkeit der Castanea sativa wird auch öfters gleich des Apfelbaumes angegeben.
Die ausgetriebene Edelkastanie ist empfindlich gegen Frost (kleiner −3 ∘C) im Frühling. Prädestiniert für späten Frühjahrsfrost sind tiefer gelegene Standorte ohne Abflussmöglichkeiten für Kaltluft und S/SW ausgerichtete Hänge, wobei durch die erhöhte Sonneneinstrahlung, die Bäume etwas früher austreiben und zusätzlich die Borke durch hohe Temperaturunterschiede am Stamm aufreißen kann. Eine weiße Stammschutzfarbe kann Stammrisse vermeiden (siehe dazu auch Abschnitt 8). Im allgemeinen treiben Bäume mit später Fruchtreifungszeit auch später aus (z.B. Sorte Brunella). Aufgrund der späten Blüte im Juni ist kein Fruchtausfall durch Spätfrost zu erwarten.
Die Eiszeiten zwischen den Warmphasen haben in den letzten 600 000-800 000 Jahren etwa einen Zyklus von 100 000 Jahren. Vor 2,7 Mio Jahren bis etwa 700 000 Jahren lag die Zyklusdauer bei etwa 40 000 Jahren.
Die letzte Kaltzeit im Alpenraum, auch Würm-Eiszeit genannt, erstreckte sich über einem Zeitraum von 115 000 bis 10 000 (Beginn des Holozäns) Jahre vor heute. Die Jahresmitteltemperaturen betrugen im Alpenvorland unter −3 ∘C (heute +7 ∘C). In dieser Phase des Temperaturminimums herrschte im eisfreien nordwestlichen Europa eine subarktische Tundrenvegetation, die aus Zwergstrauchgesellschaften, sowie einer alpinen Steinschutt- und Rasenvegetation bestand.
Auch Südeuropa war während der Zeit der maximalen Eisausdehnung vor etwa 20 000 Jahren in großen Teilen waldlos mit mit weit zerstreuten, voneinander isolierten Gehölzvorkommen an begünstigten Stellen [Wik16].
Diese Kälteperiode hatte natürlich auch desaströse Auswirkungen auf den Baumbestand der Edelkastanie in Europa. Es wurde früher angenommen, dass sich die Edelkastanie bis an das Gebiet um das schwarze Meer zurückgezogen hat. Pollenanalysen deuten mit großer Wahrscheinlichkeit darauf hin, dass die Edelkastanie auch in Refugien in Nordspanien, Mittel- und Süditalien die letzte Eiszeit überdauerte [Pat03],[CT04].
Die Zeit nach Christi Geburt ist zunächst durch eine mittelalterliche Warmzeit, welche um 900 begonnen und um 1400 endete, charakterisiert. Der wärmste Zeitraum für diese Warmzeit lag in der nördlichen Hemisphäre zwischen 950 und 1250. Danach folgte von Anfang des 15. Jahrhunderts bis in das 19. Jahrhundert hinein eine Periode relativen kühlen Klimas – die sogenannte kleine Eiszeit – wobei von 1570 bis 1630 und von 1675 bis 1715 besonders kalte Zeitabschnitte auftraten.
Klimatische Rekorde nach Christi Geburt in Europa:
Der wahrscheinlich härteste Winter in Europa nach C. war 763/764. Byzantinische Quellen wie „Die Chronik des Theophanes“ überliefern, dass das Schwarze Meer 100 Meilen von der Nordküste ausgehend gefroren war und im Februar 764 ein großer Eisstoß im Bosporus einiges zerstörte. Nach der französischen Chronik Moissiacense, ließ die Kälte viele Oliven- und Feigenbäume am Südbalkan absterben.
Der kälteste Winter Europas danach war vermutlich der von 1607/1608. Alle Flüsse, sowie die Ostsee und Bodensee froren vollständig zu. Besonders strenge Winter des 20. Jahrhunderts in Europa waren die von 1939/1940, 1940/1941, 1941/1942 und 1946/1947 und der Winter von 1962/1963 war für ganz Europa der härteste Winter des 20. Jahrhunderts. Für Mittel- und Westeuropa bemerkenswert war seine ungewöhnlich lange Frostdauer.
Anmerkungswert ist auch der Ausbruch des Vulkans Tambora im heutigen Indonesien im April 1815. Das eruptiv ausgeworfene Material bewirkte Klimaveränderungen in Nordamerika und Europa, welches dem Jahr 1816 die Bezeichnung Jahr ohne Sommer einbrachte. In Teilen der nördlichen Hemisphäre kam es durch Missernten zur schlimmsten Hungersnot des 19. Jahrhunderts.
Wetterrekorde um und in Wien:
Am 7.1.1985 wurden −19,2 ∘C in Wien und −22,1 ∘C in St. Pölten in Niederösterreich gemessen. Weitere Kälterekorde in Wien, Niederösterreich und ganz Europa: im 19. Bezirk in Wien wurden am 11.2.1929 −25,8 ∘C und am 24.1.1942 im 14. Bez. −29.1 ∘C gemessen. Es gedeihen in Niederösterreich etliche Bäume, die älter als 90 Jahre sind [Ros23].
Erste Temperaturmessungen:
Das erste gegen den äußeren Luftdruck abgedichtete Thermometer mit Skala wurde angeblich auf Anregung von Ferdinand II de’ Medici Großherzog der Toskana 1641 – das kleine Florentinische Thermometer – entwickelt. In Wirklichkeit dürfte die Entwicklung auf Galileo Galilei (1564-1642), der das Thermoskop (noch luftdruckabhängig) entwickelte, zurückgehen. Galileo Galiliei war ab 1610 als Hofmathematiker und -philosoph bei Ferdinand II tätig. Jedenfalls haben die Medici ein Wetterstationsnetzwerk aufgebaut, es umfasste von 1654-1667 11 Wetterstationen: 7 Stationen in Italien, mit den zwei Hauptstationen Florenz und Vallombrosa und vier außerhalb Italiens mit Innsbruck, Warschau, Osnabrück und Paris. Alle Stationen waren mit identischen Thermometern ausgestattet. Die beiden Hauptstationen Florenz und Vallombrosa wurden von 1654-1670 betrieben.
Die erste professionelle und systematische Erfassung von Wetterdaten begann 1781 durch die Societas Meteorologica Palatina am Observatorium Hohenpeißenberg in Bayern, Deutschland.
| Die Wurzeln, der Stamm, die Triebe, die Blattknospen und die Blütenknospen haben eine unterschiedliche Kälteresistenz. Das Wurzelsystem der Edelkastanie hat angeblich eine Frostbeständigkeit bis zu −6 (−9) ∘C . Frühjahrstriebe der Bäume haben kurzzeitig eine Frostbeständigkeit von etwa −3 ∘C . Der überwinternde Großbaum bis etwa −20 ∘C . |
Die Winterfrostbeständigkeit ist nicht nur von der absoluten Temperatur abhängig, sondern auch z.B. vom Temperaturgradienten – ein Temperatursturz ist schlechter für einen Baum, als eine langsamer Übergang zu winterlichen Temperaturen, da der Baum vor dem Winter eine Abhärtungsphase benötigt.
Die vorgelagerte Abhärtungsphase in Spätherbst wird durch kürzere Tage und niedrige Temperaturen eingeleitet, der Erfolg hängt auch an der Reduktion von Bodenfeuchtigkeit und einer geringen Stickstoffverfügbarkeit ab.
Im wesentlichen sind es drei Maßnahmen von winterharten Pflanzen, die sie vor Frost schützen:
Mythen:
Statt irgendwelche Spezialdünger nach der Wachstumperiode aufzubringen, ist es besser folgende Maßnahmen zu setzen: Als Wurzelschutz kann man Herbstlaub mit etwas trockener Erde anmischen, dieses Gemisch gut gießen und nochmals durchmischen. Dieses feuchte Gemisch dann in den Boden oberflächlich um die Bäume auftragen bzw. leicht einarbeiten. Damit bleibt das Laub auch bei Wind als Auflage fixiert. Beim Verrottungsvorgang wird durch chemische Reaktionen, welche durch Mikroorganismen und Mykorrhizen (Pilze) angetrieben werden, Wärme gebildet und die in Lösung gegangenen Huminsäuren senken auch den Gefrierpunkt des Bodens. Außerdem ist Laub ein gutes Nahrungsmittel für Würmer, die durch Krümmelbildungen und Röhrengänge die Erde auflockern. Diese lockern den Boden auch unterhalb der Laubschicht auf ((Abbildung 13)!
Abbildung 13: feuchte Laubmischung li, in den Boden leicht eingearbeitet re
Richtig schwer verrottbares Laub, wie von Nußbäumen, ist zu vermeiden. Eichen– oder Edelkastanienblätter sollten bevorzugt verwendet werden – diese sind auch nicht leicht verrottbar – bringen jedoch Huminsäuren in den Boden ein und senken damit den pH-Wert des Bodens.
Bei Jungbäumen lassen sich Frostrisse durch Sonneneinstrahlung durch Anbringung von Schilfrohrmatten oder Weißanstrichen (z.B. mit Arboflex [Jes15]) meist vermeiden.
Die chinesische und japanische Edelkastanie (C. mollissima, C. crenata) hat eine geringere Frostbeständigkeit als die europäische oder amerikanische Edelkastanie (C. sativa, C. dentata) und daher im allgemeinen auch eine geringere Frostbeständigkeit bei Einkreuzungen von C. sativa x crenata.
Vergleiche der labormässigen Bestimmung der Frostbeständigkeit in Relation zu Beobachtungen im Freien für die amerikanische Edelkastanie (C. dentata) und deren Rückkreuzungen findet man in [GGHS11]. In dem Artikel werden Angaben zur mittleren Zellmortalitätsrate Tm von terminalen Trieben gemacht. Die Temperatur Tm ist eine Schätzung, bei der 50 % aller Zellen abgestorben sind. Gemessen wurde mit der REL-Methode – siehe dazu auch Abschnitt 9.7. Zusammenfassend hat sich gezeigt, dass die labormässigen Untersuchungen für Triebe im Winter 2007 für C. dentata / C. dentata–Rückkreuzungen eine Tm von etwa −35∘C / −32 ∘C erbrachten. Werden die Triebe im Frühjahr entnommen und wieder einer Kältekammer zugeführt, so steigt die Tm auf etwa −15 ∘C für C. dentata und C. dentata-Rückkreuzungen. Die im Winter 2006–2007 im Freien aufgetretenen Frostschäden an Trieben bei Kurzzeit–Kälteminima zwischen −25∘C und nahe −30 ∘C, zeigen für C. dentata / C. dentata–Rückkreuzugen etwa ein 30 %ges / 60 %ges Absterben der Triebspitzen. Eichen (Quercus ruba) und Ahorne (Acer saccharum) zeigten keine Frostschäden.
Im Folgenden wird von einem ausgeglichenen Nährstoffgehalt des Bodens der Klasse C ausgegangen. Wurde der Boden/Rasen jahrelang mit Kompost gedüngt, dann ist genug Phosphat und Kalium im Boden vorhanden und es ist eventuell Stickstoff nachzuliefern. Als Dünger wird dann vorerst ein Volldünger mit erhöhtem N-Anteil eingesetzt werden. Langzeitdünger sollten in der Wachstumsperiode von April bis zur Mitte August in Mitteleuropa in mindestens zwei Tranchen ausgebracht werden. Bei Großbäumen wird der Dünger an der Bodenoberfläche in 0,5 m Abstand vom Stamm bis 2–3 m hinaus im Umkreis aufgestreut. Im Herbst sollte nicht mehr mit Stickstoff gedüngt werden, da ein später Austrieb die Frostresistenz des Baumes herabsetzt.
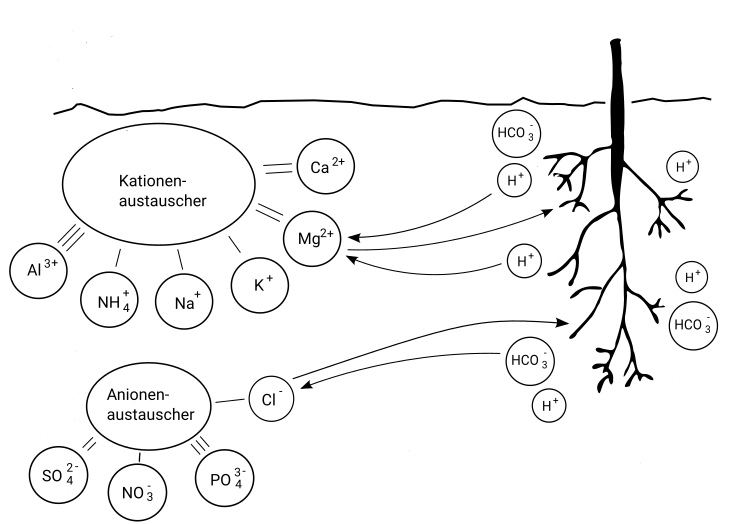
| Ist der Boden alkalisch (hoher pH-Wert), kann nur mehr wenig Stickstoff und auch Eisen von der Edelkastanie aufgenommen werden. Bei sauren Böden (niedriger pH-Wert) funktioniert die Nitrifizierung nur mehr stark eingeschränkt und die Urease ist auch vom Optimum entfernt. Edelkastanien können jedoch wie auch andere Baumarten Ammonium ohne Nitrifizierung direkt aus dem Boden aufnehmen (Abschnitt 17.5.1). Als mineralischer Stickstoffträger ist demnach Ammoniumnitrat oder Ammoniumsulfat eine gute Option. Ammoniumnitrat senkt den pH-Wert im Substrat geringfügig. Ammoniumsulfat senkt den pH-Wert stark. |
Die Düngereinbringung nach Baumdurchmesser und bei Vollertrag, dient als grobe Abschätzung und werden hier angelehnt an [Uni12] und [Viv] aufgestellt. Diese Abschätzungen ersetzen keine Bodenanalyse oder weitergehend eine Blattanalyse! Selbst wenn man die jährlich abgeführte Biomasse wie Gehäuse, Nüsse und Laub und noch zusätzlich den Holzzuwachs eruieren kann, sind die Nitratverluste im Boden nicht abwägbar.
Für eine Düngung mit einem (18/6/7/8+2) Vollwertdünger ergibt sich für einen dreijährigen Baum
20 · ta · 100 / AN = 20 · 3 · 100 / 18 = 330 g Düngermenge.
Laut [Viv] kann man für die Düngerkomponenten Ammoniumsulfat/Superphosphat/Kaliumphospat + Magnesiumoxid: 0,8/0,5/0,4 + 0,2 kg pro Baum ansetzen. Verwendet man einen Mehrstoffdünger so soll man 2–3 kg (15/9/15+2) pro Baum ausbringen.
Sind die Blätter hellgelb oder hellgrün ist wahrscheinlich mehr Stickstoffdünger notwendig.
Zweitens liefert der Jahresaustrieb auch eine Abschätzung für die Einbringung von Stickstoff. Die Länge des
Jahresaustriebs bei einem tragenden Baum sollte zwischen 15 cm und 30 cm liegen. Wenn das Wachstum mehr als 30 cm beträgt, so ist der Stickstoffdünger zu
reduzieren. Wenn das Wachstum weniger als 15 cm beträgt, ist wahrscheinlich mehr Stickstoff
notwendig [Uni12].
Es spricht jedoch nichts dagegen, wenn man einer nährstoffarmen sauren Anzuchterde gleich etwa 3 g (ist in etwa ein gestrichener Teelöffel) Langzeitdünger mit Ammoniumnitrat wie z.B. Azaleen- oder Rhododendrondünger pro 2 l Anzuchterde einmischt.
Statt Flüssigkeitsdünger, geeignet für Azaleen oder Rhododendren kaufen, gleich ein Nährsalz wie etwa Dehner Blütenzauber kaufen. Dieses auflösen und ins Gießwasser mischen.
Da man das benötigte Nährsalz nicht für jeden Gießvorgang wägen und auflösen will, kann man mit einer Stammlösung arbeiten. Gegossen wird im folgenden Beispiel mit 2 g
Nährsalz pro Liter Gießwasser:
Beim vorbereiteten Messbecher (Abbildung 14 li) sind es 50 g Nährsalz bis zur Markierung,
das Nährsalz im Stammlösungsbehälter (Abbildung 14 mi) mit warmen Wasser auflösen und den Behälter auf 1 l auffüllen.
Zum Gießen je 0,2 l der Stammlösung in die 5l Gießkanne geben und mit Wasser auffüllen.
Die Stammlösung beschriften, kühl stellen und spätestens nach 14 Tagen aufbrauchen!
| Die Löslichkeit und damit die Verfügbarkeit der Hauptdüngerelemente N/P/K steigt, grob abgeschätzt, von einem pH-Wert ab 3 beginnend linear an und ist in einem Bereich von 6,3 bis 6,8 am höchsten. |
Ein gutes Substrat soll ein ausgewogendes Gleichgewicht zwischen Wasserspeicherkapazität und Wasserabfluß herstellen und damit einfach den Wasserhaushalt regeln. Weiters soll es über längere Zeit strukturstabil bleiben und nicht verdichten. Es soll locker sein damit aerobe (mit Luft) chemische Prozesse im Substrat stattfinden können aber auch der Pflanze die notwendige Stabilität geben. Ein anorganischer Ionenaustauscher wie Tonmineralien oder Vermiculite sorgt für ein kräftiges Pflanzenwachstum.
Die von Anton Fruhstorfer um 1950 als Kultur-Substrat entwickelte Einheitserde soll der Vollständigkeit halber erwähnt werden. Diese enthält 60–70 % Weißtorf und 30–40 % kalkfreien Ton oder Untergrundlehm. Die Einheitserde hat eine Masse von etwa 300–400 g/l. Das Substrat wird gedämpft und ist daher weitestgehend keimfrei. Der eingesetzte Ton/Lehm sollte beim Zerteilen Krümmel bilden und nicht verschlemmen. Durch Aufkalkung wird der pH-Wert auf 5,6 bis 6,5 eingestellt. Je nach Verwendungszweck werden der Einheitserde auch Nährstoffe zugesetzt.
Baumschulen wollen mit leichten Substraten ohne Erde bei Containerpflanzen das Auslangen finden, weil tonhaltige Substrate schwer sind, Ton/Lehm in hoher Qualität nicht einfach verfügbar ist und das Substrat keimfrei möglichst sein sollte (Erdfreie Substrate – Soilless Substrate).
Typischerweise wird im professionellen Bereich meist mit erdfreien Substrat-Mischungen gearbeitet, welche Bestandteile wie Torfe, Perlite, Vermiculite, Lava, Bims, Holzfasern, Kokosfasern und/oder Rinde enthalten. Diese Nährstoffarmen Substrate werden meist mit Mineraldünger auf etwa
aufgedüngt.
Im amerikanischen Raum wird für die Anzucht von Edelkastanien eine Mischung von Torf, Perlit und Vermiculit im Verhältnis 1 : 1 : 1 empfohlen.
Zur besseren Wasseraufnahme wird auch eine Benetzungssubstanz zugefügt und oft wird auch Kalk zur pH-Wert-Erhöhung zugegeben.
Aufdüngung mit Kalk ist bei acidophilen Pflanzen wie die Edelkastanie, wenn überhaupt, nur in geringsten Mengen zu empfehlen.
Außerdem enthält Gießwasser meist genug Calciumcarbonat.
| Schnellwachsende Bäume in Töpfen gehören einmal pro Jahr umgetopft. Das Umtopfen sollte möglichst in der Ruhephase am besten vor dem Austrieb im Frühjahr durchgeführt werden. Gründe dafür sind: Weil der wachsende Wurzelstock oft keinen Raum mehr zur radialen Ausbreitung der Wurzeln hat und die neuen Wurzeln nur mehr im Kreis in der Topfinnenwand wachsen können. Weil die Nährstoffe im Substrat aufgebraucht sind und Zuschlagsstoffe wie Torf ist nicht mehr strukturstabil sind und daher das Substrat verdichten. |
Die Hauptsubstratkomponente für Topfpflanzen im Erwerbsanbau ist meist Torf. Torf ist ein organisches Ablagerungsprodukt, das in Mooren entsteht. Torfmoos (Sphagnum spec.) wirkt antiseptisch und kann bis zum 25-fachen seines Eigengewichtes an Flüssigkeit aufnehmen. Daher wurde im Mittelalter frisches Sphagnum auch als Wundauflage verwendet.
Trockener Torf ist im Gegensatz zu feuchtem Torf wasserabweisend (hydrophob) und muss sorgfältig angefeuchtet werden.
Da Torf meist sehr sauer ist (pHH2O-Wert etwa 4,0 bzw. pHCaCl2-Wert von etwa 3,5), ist es je nach Anwendung notwendig, diesen anzuheben. Da Torf meist mit anderen Substratkomponenten wie Kompost für Edelkastanien gemischt wird, ist von einer Anhebung mit Kalk abzuraten, da die weiteren Zuschlagsstoffe meist einen höheren pH−Wert aufweisen.
Will man unbedingt Aufkalken, so sind folgende Dinge zu beachten: Gelöschter Kalk Ca(OH)2 wirkt zu rasch und ist daher nicht zu empfehlen. Zur Aufkalkung wird meist feiner Gartenkalk verwendet. Dieser enthält z.B. 70 % CaCO3 , 25 % MgCO3 mit insgesamt 50 % Neutralisationsanteil. Das angemischte Substrat sollte auf einen Ziel-pHH2O-Wert für Edelkastanien von 5,5–6 angemischt werden. Was nicht ganz einfach ist, da auch bei angefeuchtetem Substrat der kohlensaure Kalk seine Wirkung erst nach etwa 2 Wochen voll entfalten kann und dieser dann auch den pH-Wert dann nicht mehr weiter erhöht [Hor15]. Außerdem wird durch die Zufuhr von hartem Gießwasser der pH-Wert auch angehoben.
Eine pH-Wert-Messung unmittelbar nach dem Zusammenmischen ist daher weniger sinnvoll. Es ist daher besser vorab mehrere Testmischungen zu machen, diese
anzufeuchten und mindestens 2 Wochen mit den Messungen zu warten. Grober Gartenkalk löst sich langsamer als feiner auf!
Typischer schwach bis mäßig zersetzter Weißtorf (H3–H5) enthält an Nauptnährstoffen etwa an Gesamtstickstoff: 25 mg/l, Phosphat (P2O5): 30mg/l, Kaliumoxid (K2O): 40mg/l und liegt damit ganz grob gesehen bei einem Nährstoffgehalt von einem 1/8–1/20 von normaler Blumenerde.
Für die Torf Gegner: es wird hier zur Vermehrung/Anzucht und Aufzucht nur relativ wenig Torf benötigt. Es wird 2015 noch immer Torf in großen Mengen in kalorischen Kraftwerken verbrannt!
| Mengenangaben zur Aufkalkung von Torf kann man eigentlich nicht machen, da Torf zwar oft mit einem pHCaCl2-Wert von 3,5 deklariert wird, letztendlich liegt die reale Schwankungsbreite des pHCaCl2-Wertes meist bei 2,5 bis 4,0. Je höher der Zersetzungsgrad des Torfes ist, desto höher ist der pH-Wert. Neben dem Einfluss des Zersetzungsgrades haben auch die verschiedenen Arten einen Einfluss auf den pH-Wert: Braunes Torfmoos (Sphagnum fuscum) wächst auf den kleinen Hügeln und hat einem pH–Wert von 3,5 (pHCaCl2 ??). Diese Spezies ist auch für Substrate das beste Torfmoos, weil es eine hohe Wasserabsorption besitzt und nur langsam zersetzt wird. Auch Magellans Torfmoos (Sphagnum megellanicum) nimmt Wasser gut auf, zersetzt sich jedoch schnell. Das Schmalblättriges Torfmoos (Sphagnum angustifolium), welches in Mulden wächst, hat einen pH-Wert von 4 bis 5. Auch Beiwuchs wie etwa Sumpfbinsen beeinflusst den pH-Wert [Red05]. Mittlerweile rate ich vom Aufkalken von Torf für Edelkastanien ab, da genug Calciumkarbonat durch das Gießwasser ins Substrat eingebracht wird! |
Ton besteht im wesentlichen aus Kaolinit Al2O3· 2SiO2· 2H2O, dem quellbaren Montmorillonit Al2O3+4SiO2+H2O+nH2O und MgO und CaO. Montmorillonit kann bis zu die siebenfache Wassermenge binden! Daneben finden sich in den gewöhnlich braunen oder gelblichen Tonen wechselnde Mengen von Fe-, Mn-, Mg-, Ti-, P-, N- usw. Verbindungen, Humusbestandteile, Spurenelemente, die teils chemisch gebunden, teils physikalisch an die sehr kleinen Tonteilchen absorbiert sind.
Sind Feldspat, Glimmer und andere aluminiumartige Doppelsilikate für lange Zeit Wasser ausgesetzt, so verwittern sie zu Ton. Reiner Ton hat normalerweise einen recht hohen pH-Wert von bis über 9. Quellbare Tone puffern Nährsalze durch ihre hohe KAK (siehe auch Abschnitt 17.2.2). Außerdem stabilisieren Tone durch sein hohes spezifisches Gewicht (trocken etwa 2 kg/l) Pflanzen im Topf.
Mischt man Ton z.B. mit Torf in das Substrat, so wird sich das Gemisch zu einer kompakten Masse verklumpen und dadurch wenig Luft im Substrat verbleiben
und damit auch der Wasserabfluss behindert werden. Bei sandigem Lehm bzw. Löss bleibt das Substrat bleibt bröselig.
Es gibt auch mehr oder weniger kalkstein-freien Lehm und Löss. Ein ganz niedriger pH-Wert kann bei tonigen Böden toxische Konzentrationen von Aluminium freisetzen!
Lehm ist im Wesentlichen von den Bruchstückgrößen her eine Mischung aus Sand (Korngröße > 63 µ m), Schluff (Korngröße > 2 µ m) und Ton (Korngröße < 2 µ m). Ziegellehm besteht je nach Anwendung (Vollziegel, Lochziegel, Dachziegel) aus 20 bis 60% Ton, feinem Sand (großteils Quarzsand SiO2) und bis zu 25 % Calziumcarbonat/Kalksteinsand (CaCO3) [Thi16]. Lehm ist wie Ton ein guter Ionenaustauscher und daher eine nicht unwesentliche anorganische Komponente von Substraten.
Löss ist hauptsächlich eine Mischung von kalkhaltigem Schluff, Ton (5 bis 20 %) und Feinsand. Mineralogisch besteht Löss zum größten Teil (50 bis 80 %) aus Quarzkörnern mit 8 bis 20 % kalkigen Bruchstücken. Löss ohne Kalkstein wird als sandiger Lehm bezeichnet. Feuchter Löss lässt sich im Gegensatz zu Ton oder Lehm nicht mehr zwischen den Fingern kneten.
Für Lehm, Löss lässt sich mit dem Zuwachsen der Korngröße ein gleichsinnige Erhöhung des Anteils von Quarzsand SiO2 erwarten.
| Achtung: Ton, Lehm oder Löss kann hohe Anteile von Calciumkarbonat enthalten. |
Abbildung 15: Ziegellehm li, Löss re
Vermiculite sind glimmerähnliche Tonsilikate, welche chemisch in die Gruppe der Schichtsilikate eingeordnet werden. Expandierte Vermiculite wurden gebläht, sind daher sehr leicht und haben außerdem die Fähigkeit viel Wasser zu speichern.
Da Vermiculit keimfrei und nahezu pH-neutral ist, eignet es sich hervorragend für die Aufzucht vieler Pflanzen bei Düngerzusatz. Vermiculit kann auf Grund seiner Struktur viele Kationen (hohe KAK) binden. Expandierte Vermiculite sind jedoch in Substraten nicht lange formstabil - sie fallen nach einigen Monaten wieder zu Plättchen zusammen und zerbröseln.
Grobe Rindenschnitzel als Zuschlagsstoff können das Substrat auflockern. Frische Rindenstückchen, besonders Nadelholzrinde oder auch grobe Holzspäne bleiben lange strukturstabil und werden nur langsam abgebaut. Für Jungpflanzen sind diese Zuschlagsstoffe ungeeignet, wenn diese noch nach Harz riechen, da sie auf die Wurzeln phytotoxisch wirken. Wahrscheinlich sind die enthaltenen Phenole Terpene (hier Gemische von Harz und ätherischen Ölen), Tanninsäuren und auch das Mangan phytotoxisch.
Erfolgt ein anerober (ohne Sauerstoff) Abbau der Rinde/Holz im Substrat, so wird neben Stoffen wie Methan auch Essigsäure gebildet und der pH-Wert kann auf bis etwa 2,5 hinunter sinken! Wird die Rinde/Holz im Substrat aerob abgebaut, so hebt das abgegebene Calcium den pH-Wert.
Generell sollten Hartholzrinden und Holzfasern gut vor-kompostiert werden, da dadurch die Substratkomponenten ihre Toxizität verlieren, Wasser besser aufnehmen können und auch mineralisierte Nährstoffe wie Stickstoff und Calcium abgeben. Bei normaler Kompostierung wird im Wesentlichen organisches Material (Kohlenstoffverbindungen) mit Sauerstoff (aerober Prozess) und Wasser unter Hilfe von Bakterien, Aktinomyceten und niederen Pilzen zu Huminstoffen, Kohlendioxid, Methan, Kohlenmonoxid, Wasser und zu wasserlöslichen Mineralstoffen abgebaut. Bei diesem Prozess wird Wärme produziert. Kompost erreicht bei einer Heißrotte durch Eigenerwärmung etwa 60−70 ∘C.
Wird Kompost mehrheitlich aus Ästen und wenig anderen Komponenten wie Gras und Küchenabfällen im Schnellverfahren gewonnen, weist dieser einen hohen pH-Wert auf. Im Kompostwerk der Lobau, dauert die Rotte 8 bis 10 Wochen. In der thermophilen Phase (Heißrotte) steigt die Temperatur durch Bakterien in der Miete auf maximal 60°C bis 70°C. Durch die hohe Temperatur sterben die meisten schädlichen Baktierien und die Samen verlieren ihre Keimfähigkeit.
Kompost aus dem Kompostwerk Lobau wird an den Wiener MA48 Mistplätzen netterweise in Kleinmengen kostenfrei abgegeben. Gemessen wurde von mir im April 2019 ein pHH2O-Wert von 8,3. Von diesem Kompost ist für säureliebende Pflanzen als Zuschlagstoff abzuraten!
Bei einem sogenannten Wurmkompost, gebildet durch Eigenkompostierung von Küchenabfällen, Gras und dünnem Astwerk in einer Box im Garten, wurde ein pHH2O-Wert von etwa 7,5 gemessen. Die Reifezeit lag bei einem Jahr.
Kompostisierter Stallmist von Wiederkäuern, ist ein nährstoffreicher Zuschlagsstoff. Bei Stallmist, lange durch Würmer aufbereitet, liegt der pHH2O-Wert bei etwa 7,5. Für Nährstoffanteile von Stallmist siehe Abschnitt 17.8.
Die Faserschicht wird aus der Kernumhüllung der Nuß – eigentlich eine Steinfrucht – der Kokosnusspalme (Cocos nucifera) gewonnen. Das Fasermaterial bleibt lange strukturstabil und kann viel Wasser speichern. Kokostorf oder Kokohumus oder auch Kokoserde genannt wird aus zerkleinerten, zerriebenen Fasern erzeugt. Kokostorf ist im trockenen Zustand im Gegensatz zu Torf hydrophil und ist ein guter Kationenaustauscher. Kokostorf hat einen hohen Zellulose- und Lignin-Gehalt aber nur einen geringen Nährstoffgehalt. Kokoschips bestehen aus geschnittenen Kokosfasern und aus Teilen der Außenhülle (Exocarp) der Nuß. Als Zuschlagsstoff verwendet hält es das Substrat lange locker.
Die Firma FibreDust Europe BV bietet zu Ziegeln gepresste Kokoschips an. Bei diesem Material habe ich einen pHH2O-Wert von 5,9 gemessen.
Anmerkung: Minderwertig verarbeitete Kokosfasern kann einen hohen Salzgehalt aufweisen und muss daher gewaschen werden. Angeblich wird das Palmmaterial auch manchmal im Meer gewässert.
Frische Holzkohle und auch Asche bildet mit Wasser zunächst Laugen (Aschenlauge war früher ein Reinigungsmittel, pH-Wert bis 12). In Wasser gelöste Holzkohle oder Asche bleibt auch unter Kohlendioxidzufuhr (CO2) leicht basisch. Diese Zuschlagsstoffe gehören nicht unbehandelt in das fertige Substrat. Angeblich kann man Holzkohle mit Zitronensäure so neutralisieren, dass sie rasch wurzelverträglich und auch mikrobenverträglich im Komposthaufen ist.
Will man die Edelkastanien zur Keimung bringen oder nur kühl lagern, so kann man Torf als Keimungs-Substrat oder Lagermaterial verwenden. Hochmoortorf mit einem niedrigen pHCaCl2–Wert von etwa 3,5 ist meist ausreichend keimfrei.
Möchte man einen Edelkastanienbaum aus einer Nuss selbst ziehen oder eine Ammenveredelung durchführen, so sollte man ein Substrat verwenden, das Wasser speichern kann, locker ist, damit Gaszirkulation erlaubt, den Wurzeln halt bietet, eventuell durch das Gewicht den Topf stabilisiert und möglichst wenig mit unerwünschte Pilzen, Parasiten und keimbaren Samen durchsetzt ist.
Anzucht-Substrat mit Torf und Vermiculit
Als erdfreies und nahezu keimfreies Kurzzeitsubstrat, wie es für eine Ammenveredelung notwendig ist, bietet sich Torf und Vermiculit als Zuschlagsstoff an.
Das Volumenmischungsverhältnis von Torf : Vermiculit kann mit etwa 3:1 v/v (Volumenverhältnis) angesetzt werden. Der Vermiculit wird mit im Wasser aufgelösten chloridarmen Nährsalz aufgedüngt, sodass auf das Fertigsubstrat etwa 1 g /l Nährsalz kommt. Der aufgedüngte Vermiculit wird dann in den Torf gemischt.
Beispielhaft wird hier Substrat-Anmischung von etwa 40 l Fertig-Substrat unter Verwendung von 2 Eurokisten E3 (Innenmaße etwa: 56x36x29 cm) angeführt. In eine Eurokiste 15 cm hoch feinen Torf – eventuell durch ein Sieb mit 6 mm Maschenweite sieben – einschichten. Den Torf leicht gießen und durch mischen gleichmäßig durchfeuchten. In die zweite Kiste etwa 5 cm Vermiculit wie Isola Vermiculite M mit Körnung 2–4 mm oder auch etwas gröber einschichten.
40 g Nährsalz wie etwa Dehner Blütenzauber oder Compo Blumenprofi oder Compo Hakaphos blau[Dü15] oder Hauert Planaktiv (Typ A) in
einem Liter warmen Wasser auflösen (siehe auch Abschnitt 17.9).
Diese Nährsalzlösung in eine größere Gießkanne mit Brausemundstück geben und nochmals mit Wasser verdünnen und gleichmäßig über den Vermiculit gießen und
gleichzeitig den Vermiculit vorsichtig durchmischen – dabei die Vermiculitstückchen nicht zusammenpressen.
Einen Teil des Torfes in eine Hälfte von Mischkiste 2 geben und den nicht abgedeckten Vermiculit aus 2 in die 1 geben. Den Inhalt beider Kisten gut
durchmischen, dann eine Kiste in die andere Kiste entleeren und das Substrat nochmals mischen bzw. nach-feuchten und mindestens 14 Tage vor Verwendung stehen lassen.
Testweise Aufkalkung von Torf:
| pHCaCl2? | pHH2O | |
| Immergrün Torf (ImT) deklariert | 2,5–4,0 | 4,0–4,7 |
| Gemessen ImT | ≈ 4,2 | |
| Substrat (ImT + Verm. + Kalk); 14 Tage | ≈ 6,3 | |
Abbildung 16: Anzucht Substrat: Torf + Vermiculit li, Mischung re
Abbildung 17: Eurobox E3 als Mischkiste
Es gibt viele gute Substrate für die Edelkastanien, solange der Kalkgehalt gering bleibt. Auch sandiger Lehm ist eine gute Substratkomponente, solange der Kalkanteil gering ist!
Als einfache Variante bietet sich fertig angemischte und vorgedüngte Rhododendron-erde als Substrat an. In das Substrat mischt man noch etwa 10 % Ziegelsplitt mit einer Körnung etwa 8 bis 16 mm. Den Splitt kann man z.B. aus kaputten unglasierten Tontöpfen gewinnen. Den Topf in ein Tuch einwickeln und mit einem Hammer auf einer harten Unterlage zerschlagen – trotzdem Schutzbrille tragen.
Die Rhododendron-erde ist sauer, bei einem pHCaCl2–Wert zwischen 4 bis 5. Die Scherben bilden Hohlräume im Substrat und binden auch etwas Wasser und Nährstoffe.
Hier ein Rezept für ein lockeres Substrat, welches für eine Wachstumsperiode stabil bleibt, das heißt der Pflanze einen guten Halt gibt und außerdem einen guten Nährstoffpuffer besitzt. Dieses besteht aus den Komponenten Torf, Kokosfasern, die sich nur langsam zersetzen. Weiters aus vorgedüngtem Vermiculit in der Körnung 2–4 mm als mineralischer Ionenaustauscher. Zur mechanischen Lavakies in der Größe 8–16 mm und Fichtennadeln.
Ungefähres Mischungsverhältnis der Grundkomponenten (v/v):
Torf: Kokosfasern : Fichtennadeln : Vermiculit : Lavakies = 8 : 1 : 1 : 1/2 : 1/2
Vorbereitung der Komponenten: Mehrere Jahre abgelegene trockene Fichtennadeln oder andere kürzere Nadeln aufsammeln. Frischen Nadeln sind wegen der darin, enthaltenene Phenole und auch Terpene ungeeignet. Der Torf soll locker sein. Sollten sehr grobe Stücke im Torf sein, diesen mit einem Sieb 6 bis 10 mm Maschenweite vorsieben. Möglichst roten Lavakies in der Körnung 8 bis 16 mm besorgen. Die rote Farbe deutet auf einen höheren Eisengehalt hin. Kokosfasern gibt es als Briketts, die man vorab aufquellen läßt.
Der Vermiculit wird mit in Wasser aufgelöstem Nährsalz wie Dehner Blütenzauber aufgedüngt, sodass das Fertigsubstrat einen Nährsalzgehalt von etwa 3 g/l aufweist. Bei einem Liter Vermiculit sind 30g aufgelöstes Düngersalz anzusetzen.
Mischung:
Die feuchten Kokosfasern in den Torf mischen, danach den aufgedüngten Vermiculit vorsichtig druntermischen und die Mischung gießen. Das unfertige Substrat aber nur soweit gießen, dass sich Wasser mit der Hand nur schwer herauspressen läßt. Danach die trockenen Fichtennadeln und den Lavakies beimischen.
Empfehlenswert ist auch die Zugabe von organischem Dünger wie Biosol in einer Dosierung von etwa 3 g/l. Etwas Biosol kann nämlich die Grundlage für ein gutes Mykorrhiza-Wachstum sein.
Möchte man Mischung vereinfachen, so sollte man jedenfalls die Komponenten Torf, Kokosfasern und Fichetennadeln verwenden. Die Kokosfasern sollte man dann gezielt aufdüngen.
Das Volumenmischungsverhältnis (v/v) von lockerem Torf : Vermiculit : Lavakies wurde hier mit 4 : 1 : 1/2 angesetzt.
Weist der Torf sehr grobe Komponenten auf, ist dieser dieser vorab zu sieben (6–10 mm).
Der Vermiculit wurde mit in Wasser aufgelöstem Nährsalz wie Dehner Blütenzauber aufgedüngt, sodass das Fertigsubstrat einen Nährsalzgehalt von etwa 3 g/l aufweist. Der aufgedüngte Vermiculit wurde dann in den Torf gemischt und angefeuchtet. Der Lavakies wurde aus Gewichtsgründen erst beim Eintopfen zugegeben.
In Torf etwas Lavasand mit einer Körnung bis 3 mm einmischen, dann gesiebten Grünschnittkompost (gesiebt mit etwa 6 mm Maschenweite) dazugeben, kompostierten Stallmist von Wiederkäuern und zuletzt Lavakies in einer Körnung 8 bis 16 mm im Substrat verteilen. Aus Gewichtsgründen den Lavakies vielleicht unmittelbar vor dem Eintopfen einmischen.
ungefähres Mischungsverhältnis der Grundkomponenten (v/v) als Basis:
Torf : Lavasand : Grünschnittkompost : kompostierter Stallmist : Lavakies = 5 : 1 : 1 : 2 : 1
Empfehlenswert ist auch die Zugabe von organischem Dünger wie Biosol in einer Dosierung von etwa 3 g/l.
Allgemeine Hinweise für selbsthergestelle Spezialsubstrate: Mindestens 50 % Torf verwenden damit der pH-Wert unten bleibt. Grünschnittkompost, möglichst Wurmkompost verwenden, da grober Holzkompost einen recht hohen pH-Wert aufweisen kann (siehe dazu auch Abschnitt 5.1.5)! Frischer Stallmist ist ungeeignet. Für Topfpflanzen muss das Substrat wurm-frei sein! Wenn Kompost eine richtige Heissrotte durchlaufen hat, dann ist er auch wurm-frei! Im Zweifelsfall den kompostierten Stallmist z.B. mit einem alten Mikrowellenherd erhitzen. Als Richtwert gibt: 5l im Plastiksack brauchen etwa 12 Minuten um eine Temperatur von 70 ∘C zu ereichen. Braucht man nur Kleinmengen, so kann man auch pelletierten Rinderdung als Ersatz für den Stallmistkompost verwenden – dann etwa einen Teil statt zwei Teile beimischen. Für einen hohen Luftanteil im Substrat ist der faserige Torf und auch der Lavakies zuständig. Keinen minderwertigen Lavakies verwenden, welcher einen nennenswerten Anteil an schwerem Basalt ohne Poren enthält.
Die Basismischung etwas anfeuchten und vor dem Gebrauch etwa 2 Wochen stehen lassen!
Bei der Baumpflanzung dem Aushub etwa 20 % kompostierten Stallmist und eventuell eine handvoll pelletierten organischen Dünger wie Biosol zugeben, die Komponenten gut durchmischen und das Pflanzloch um und unter dem Baum damit auffüllen.
Als Mykorrhiza bezeichnet man eine Symbiose von Pilzen und Pflanzen, bei der ein Pilz mit dem Feinwurzelsystem einer Pflanze in Kontakt ist. Durch den die Wurzeln umgebenden Pilzmantel wird die Wurzeloberfläche stark vergrößert. Der Partner-Pilz hilft dem Baum bei der Wasser- und Nährstoffaufnahme aus dem Boden. Dadurch kann die pH-Wert-Toleranz der Pflanze erhöht werden, da durch Mykorrhiza zusätzliche Nährstoffe mobilisiert werden. Besonders gut können Mykorrhiza gebundene Phosphate aufschließen und für die Pflanzen verfügbar machen. Außerdem helfen Mykorrhiza-Pilze bei der Produktion von Hormonen und wehren Krankheitserreger ab. Speziell wird der Pathogendruck z.B. durch Pythium-, Phytophtora- oder Fusarium-Arten auf die Wurzeln vermindert [EB15]. Im Gegenzug bekommt der Pilz Kohlehydrate und Vitamine von der Pflanze.
Differenziert nach Eigenschaften werden die Mykorrhizen auf drei Gruppen aufgeschlüsselt: Ekto-Mykorrhiza-Pilze
Endomykorrhiza und Arbuskuläre Mykorrhiza. Eine Aufteilung in 7 Gruppen findet man in [Gra09].
Arbuskuläre Mykorrhiza
Dies ist die häufigste Art von Mykorrhiza. 70-90 % der Höheren Landpflanzen sind mit Pilzen in Form einer arbuskulären Mykorrhiza vergesellschaftet.
Arbuskuläre Mykorrhizen sind eine besondere Form der Endomykorrhiza-Pilze. Charakteristisch ist die Bildung von Arbuskeln – das sind verzweigte, zarte Hyphen in Bäumchenform
innerhalb der Wurzelzellen. Im Wurzelgewebe der Pflanze bilden sich dickwandige Pilzzellen. Auch Bäume, die untypisch für eine Vergesellschaftung mit Arbuskulären Mykorrhizen sind, können, wie
für die Eiche gezeigt, als Keimlinge zunächst eine Vergesellschaftung ausbilden [Sch09].
Endomykorrhiza-Pilze
Hier dringt ein Teil der Hyphen (sind die fadenförmigen Zellen) des Pilzes in die Zellen der Wurzelrinde des Pflanzenpartners ein.
Symbiosepartner sind überwiegend krautige Pflanzen, nur in seltenen Fällen Bäume.
Ekto-Mykorrhiza-Pilze
Der Pilz wächst auch in die Wurzelrinde hinein, dringt aber nicht in die Wurzelzellen ein. Das Feinwurzelsystem bildet sich bei Symbiose etwas zurück und wird vom Pilz übernommen.
Für die Edelkastanie sind Ekto-Mykorrhiza-Pilze als Symbiosepilze geeignet
Die Familie der Ekto-Mykorrhiza-Pilze umfasst unter anderem Pilze wie Trüffeln, Fliegenpilze, Herrenpilze, Kaiserlinge, Grüner Knollenblätterpilze, Täublinge.
In [Gra09] wird eine Anzahl von 34 Ekto-Mykorrhiza-Pilzen als mit der Edelkastanie vergesellschaftsfähig (Castanea sativa) angegeben. In wärmeren Regionen ist der Erbsenstreuling (Pisolithus tinctorius), in kälteren Regionen auch der kahle Krempling (Paxillus involutus) einsetzbar. Weitere verwendbare Pilze sind die Milchlinge (Lactarius), Täublinge (Russula), Schleierlinge (Cortinariaceae), Stoppelpilze (Hydnaceae) und Dickröhrlinge (Boletus).
Bei Becherpilzen wie dem Erbsenstreuling werden oft nur die Pilzsporen – welche von diesem leicht zu gewinnen sind – zum Kauf angeboten. Mycelpräparate sind aufwendiger zu ziehen und sind als Produkt meist weniger als 4 Wochen haltbar! Dafür sind diese Impfpräparate mit Bäumen erfolgversprechender zu vergesellschaften.
Hohe Gaben von Mineraldüngern können die symbiotische Effekte mindern. Die Anwendung systemisch wirkender Fungizide ist nicht möglich [EB15].
Als Antagonist soll hier noch die Gruppe der Trichoderma (Saprotroph, Mycoparasit) angeführt werden:
Trichoderma
Trichoderma-Arten sind filamentöse Pilze, die im Boden, Pflanzen, in verfaulenden Pflanzenresten oder auch Holz sind. Sie sind äußerst wichtig in der
Rhizosphäre und interagieren zwischen Pflanze, anderen Mikroorganismen und Boden.
Trichoderma harzianum und Trichoderma koningii werden in vielen Pilzpräparaten in Form von Sporen angeboten.
Trichoderma harzianum bildet angeblich zahlreiche Enzyme wie z.B. Cellulase, Cellobiase, Hemicellulase, Pektinase und Glucoamylase, durch diese er pflanzen-pathogene Pilze angreifen kann.
Trichoderma harzianum soll aber auch aggressiv gegen Pilze wie dem Agaricus bisporus vorgehen!
Da die Edelkastanien Tiefwurzler sind, nimmt man ein höheres Pflanzgefäß. Generell sollten Topfpflanzen einmal pro Jahr, am besten im Frühjahr, umgetopft werden. Das Umtopfen im Schatten oder Innenraum durchführen.
Eine Tetra-packung ist als Aufzuchttopf nur bedingt geeignet, da der Karton aufweicht und daher auf Dauer nicht die notwendige Stabilität liefert. Bekannte Hersteller von viereckigen Anzucht– und Aufzuchttopfen (engl. square pots) mit seitlichen Schlitzen sind Soparco, Cep, Teku, Nuova Pasquini & Bini (12x12x20 cm – 2 l, 15x15x20 cm – 3,6 l, 18x18x23 cm – 6 l, 25x25x25,5 cm – 11 l).
Der in Abbildung 23 gezeigte eckige Aufzuchttopf mit 15x15x20 cm (3,6 l) ist für unterjährige schwachwüchsige Bäume geeignet.
Besser ist es gleich nach dem Vereinzeln der Bäume mit einem Topf mit den Abmaßen von 18x18x23 cm – 6 l zu starten.
Den Wurzelballen vorsichtig aus dem alten Topf/Kiste/Plastiksack ziehen, ausgraben bzw. ausleeren und den Wurzelstock bei älteren Topfpflanzen im Randbereich etwas auflockern. Sind die Wurzeln in der Innenwand des alten Topfes im Kreis gewachsen, dann diese mit den Fingern auflockern und wenns nicht anders geht etwas aufreißen – auf einen Wurzelschnitt aber möglichst verzichten.
Je nach Topfgröße etwa 2 bis 3 cm Ziegelsplitt oder Lavakies in der Körnung 8 bis 16 mm am Boden als Drainage einschichten. Den Topf mit der Drainage auf der Arbeitsfläche aufstoßen, damit sich der Splitt etwas verdichtet. Die Substrataufbereitung wird in Abschnitt 5 gezeigt.
und es vom zum Topfrand noch etwa 2 bis 3 cm sind.
Den Baum wieder aus dem neuen Topf herausnehmen, die Substratschicht hinzugeben und etwas angießen. Dann den Baum in den Topf stellen, ausrichten und seitlich mit Substrat auffüllen, wobei man mit den Fingern etwas nachdrückt um eventuelle Höhlräume zu entfernen.
Nun klopft man den Topf wieder einmal, zweimal auf die Arbeitsfläche und verdichtet damit das Substrat. Wenn notwendig etwas Substrat am Topfrand ergänzen.
Zum Schluss den Baum gießen und nicht mehr nachdrücken.
Es wird hier in Abbildung 23 eine Drainage aus zwei Schichten gezeigt. Die beiden unteren Schichten mit Lavakies und grober Torf (Siebreste) sollen ein rasches Ablaufen des Gießwassers ermöglichen.
Eine Abdeckung des Substrates mit Steinen, Lavakies oder Rindenmulch verhindert, dass Insekten Eier in das Substrat legen und sich damit Larven einnisten, welche sich von den Baumwurzeln ernähren. Nach dem Umtopfen den Baum ein paar Tage im Schatten oder Halbschatten stehen lassen. Generell die schwarzen Anzuchttöpfe nie der prallen Sommersonne aussetzen!
Abbildung 24: gut verwurzelte Topfpflanze [Kö15a]: 18x18x23 cm mit Lavakies als Abdeckung li, ohne Topf re
In Österreich sollten Bäume erst ab etwa 1 cm Stammdurchmesser ins Freie ausgepflanzt werden. Die beste Auspflanzzeit ist in Mitteleuropa das Frühjahr, nachdem der Bodenfrost vorüber ist. Für veredelte Bäume sollte mindestens eine Fläche von 7 x 7 m oder besser 10 x 10 m Fläche vorgesehen werden.
In Plantagen werden meist vorerst mehr Bäume als benötigt gepflanzt. Wenn Baumkronen ineinander wachsen, werden Bäume entfernt. In [Gau07] wird gezeigt, wie systematisch vorgegangen werden kann. Es werden Bäume in 10 m Reihenabstand und innerhalb einer Reihe mit 1,5 m Abstand ausgepflanzt. Die sogenannten Füllerbäume bzw. Weicherbäume werden nach 15 bis 20 Jahren Standzeit entfernt. Die verbleibenden Bäume stehen dann auf einem Raster von 9 x 10 m.
Baumpflanzungen nie in der prallen Sonne durchführen.
Wenn man noch nie einen Baum gepflanzt hat ist folgende Methode für einen kleinen Baum im Topf mit max. 2 m Höhe eine guter Ansatz: ein Pflanzloch 3-mal im Durchmesser und eineinhalb mal so tief wie der Topf groß ist graben. Das Loch mit aufbereiteter Aushuberde – wird in Abschnitt 5.9) gezeigt – bis auf untere Topfhöhe auffüllen und leicht verdichten. Nun den Topf mit der Edelkastanie in das Loch stellen – auf 3 cm unter das Original-Niveau justieren. Danach den Topf herausnehmen – nun ist auch der richtige Zeitpunkt um die Stützpfähle einzuschlagen – und dann den Boden im Pflanzloch etwas angießen.
Den Wurzelballen mit der Erde vorsichtig aus dem Topf herausziehen oder wenn nicht möglich, bei einem Kunststofftopf, vorab aufschneiden und dann erst herausziehen. Die Pflanze nie aus dem Topf reißen. Den Wurzelballen im Randbereich etwas auflockern. Sind die Wurzeln in der Innenwand des Topfes im Kreis gewachsen, dann diese mit den Fingern auflockern und wenn es nicht anders geht etwas aufreißen – auf einen Wurzelschnitt aber möglichst verzichten.
Den Baum nun in das Loch stellen, ausrichten, die Erde seitlich einbringen und andrücken, wobei der Baum nicht tiefer oder höher als im Topf stehen sollte. Eventuell zusätzlich einen kleinen Gießwall um den Baum ziehen dann wieder ausgiebig gießen. Wichtig: die Veredelungsstelle darf nicht unter der Erde liegen!
Löcher für Stützpfähle immer vorbohren bzw. Pfähle einschlagen bevor man den Baum einpflanzt! Das bedeutet auch, die Stützpfähle werden nie durch den Wurzelballen geschlagen!
In der Minimalvariante einen Stützpfahl 30−45∘ schräg in die Erde drücken – in Richtung der Hauptwindrichtung (Pfahl im LUV). Dann den Baum am Pfahl mit einer Achterschlinge festbinden, damit der Baum nicht am Pfahl scheuert.
Eine stabile Variante ist zwei Pfähle einzuschlagen, ein Querverstrebungsholz anschrauben und den Stamm z.B. mit in Streifen geschnittenen Fahrradschläuchen und Tackerklammern zu fixieren (Abbildung 25).
Abbildung 25: Baumstützung mit zwei Pfählen
Eine sehr stabile aber aufwendige Konstruktion, welche meist für größere Bäume angewendet wird, ist das Einschlagen von drei Pfählen und die Anbringung von Eckverbindern (Abbildung 27).
Um temperaturbedingte Stammrisse, wie sie im Winter bei Sonnenschein auftreten können, zu vermeiden, wird ein Weißanstrich zur Lichtreflexion empfohlen. Abbildung 27 zeigt einen Anstrich mit LX60 und Arbor-Flex [Jes15].
Abbildung 27: Vorbildliche Baumpflanzung in Schönbrunn: Dreibein, Gießmulde und Weißanstrich gegen Frostschäden
Wenn notwendig, ein Gitter gegen Wildschäden anbringen. Keine dichten Hüllen gegen Wildverbiss oder Fegeschäden verwenden.
Abbildung 29: Wildschaden bis in 1,5m Höhe
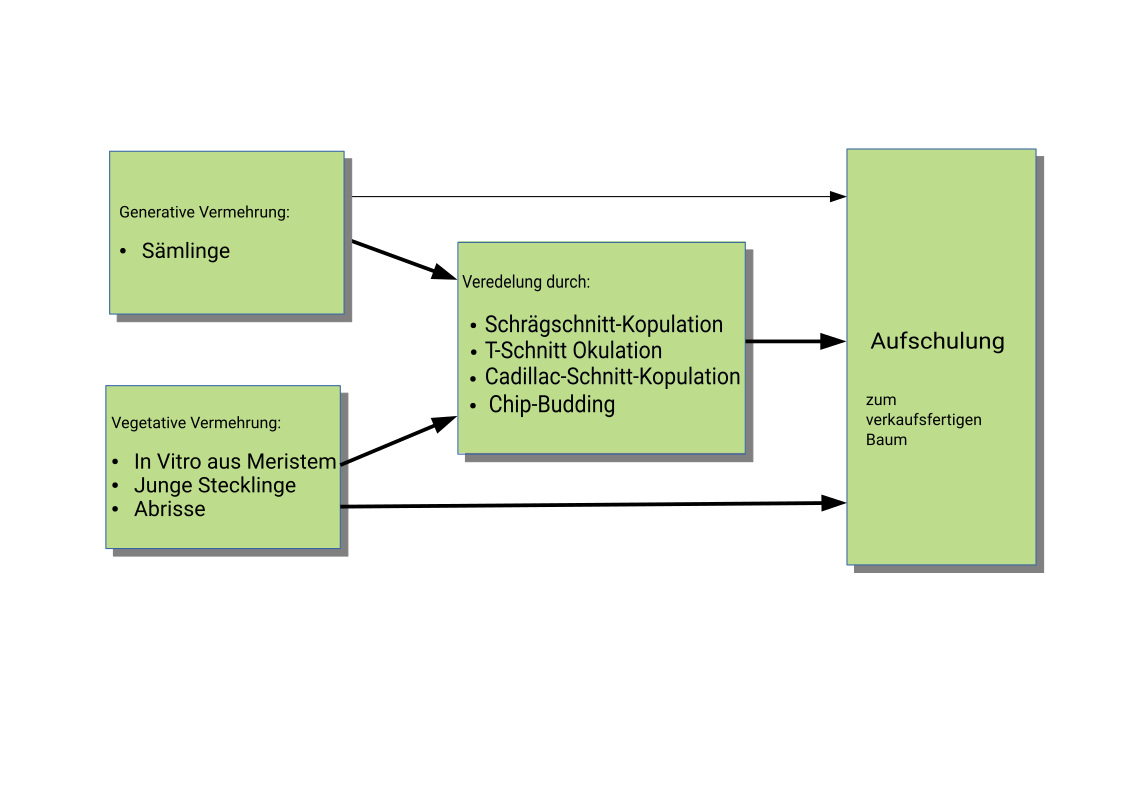
Bei Pflanzen wird grundsätzlich zwischen Vermehrung durch Samen, der sogenannten generativen Vermehrung und der Vermehrung durch Teile der Mutterpflanze, der vegetativen Vermehrung unterschieden. Bei der vegetativen Vermehrung hat die neue Pflanze das gleiche Erbgut wie die Mutterpflanze.
Alle Edelkastanienarten besitzen einen diploiden Chromosomensatz mit 12 Chromosomenpaare 2n = 24 (x = 12). Eine Genomdatenbank der Buchengewächse ist unter [oT08] zu finden.
Die erste Kindergeneration (F1) zeigt also anhand der Ausprägung (Phänotyp) eine Eigenschaft nur aus der männlichen oder weiblichen Elternpflanze und nicht ein Gemisch. Generell ist bei reinerbigen Sorten das Vermehrungsresultat in der nächsten Generation (F1) vorhersehbar. Dazu muss man allerdings wissen, welche Merkmale der Eltern dominant sind. Sind die Eltern reinerbig und auch genetisch identisch, dann sind auch alle Generationen gleich (F1 – Fn).
Bei der generativen Vermehrung von Fruchtsorten ist das Resultat nicht einfach vorhersehbar – meist sind sie nicht reinerbig, dazu müsste nämlich über viele Generationen eine Bestäubung mit der gleichen Art erfolgt sein und mischerbige Pflanzen ausgeschieden werden. Möchte man ein Merkmal beständig erhalten (homozygot – reinerbig), dann muss man nach einer Kreuzung mischerbige Eigenschaften des Merkmals herauszufiltern und diese Pflanzen für die Samenproduktion nicht verwenden.
Dieses Herausfiltern geschieht über Rückkreuzungen mit der reinerbigen (homozygot) rezessiven Elterneigenschaft. Das Kreuzungsprodukt zeigt das dominante Merkmal zu 100 %.
Es wird angegeben, dass mindestens 8 Generationen erforderlich sind um gewünschte Eigenschaftskombinationen stabil zu halten!
Ein wesentlicher Bestandteil neuer Züchtungen ist die Resistenz gegen Schädlinge. Drei Schädlinge machen der europäischen Edelkastanie (Castanea sativa) Probleme, wobei zwei aus dem asiatischen Raum eingeschleppt wurden (Abschnitt 10): der Kastanienrindenkrebs – ein Schlauchpilz, die Gallwespe und die Tintenkrankheit – Pilz (siehe auch Abschnitt 11).
Die weitgehende Resistenz gegen Kastanienrindenkrebs der Castanea crenata (jap.) und Castanea mollissima (chin.) möchte man z.B. in europäische Sorten einbringen, ohne dabei Geschmack, Größe, Schälbarkeit, Frühreife der Frucht oder die Kälteresistenz zu verlieren.
Daher ist die Erzeugung von Hybriden (Mischlingen) mit verbesserten Eigenschaften durch generative Vermehrung, ein Forschungsgebiet, dass durch den Erwerbsanbau forciert wird. Anmerkung: neben der Kreuzungen von Pflanzen um gewünschte Eigenschaften zu selektieren, gibt es die Möglichkeit durch Gentechnik Gene hinzufügen und Gene zu tauschen.
Die Verzweigung der Edelkastanie erfolgt sympodial. Laut [Wik15d] sterben die Endknospen im Herbst ab, die nächsttiefere Seitenknospe übernimmt im Frühjahr das Längenwachstum des Triebes. Anmerkung: Diese Anmerkung in Wikipedia dürfte nicht richtig sein, da ich bei in Österreich im Dezember geschnittenen Edelreisern terminale Austriebe feststellen konnte. Richtig ist vielleicht, dass die empfindlicheren Blütenknospen im Winter absterben!
Es befinden sich bei der Edelkastanie männliche und weibliche Blüten auf dem gleichen Baum – Einhäusigkeit (Monözie). Trotzdem gehört die Edelkastanie zu den Arten, die sich nicht selbst befruchten können und ist deshalb auf Fremdbestäubung angewiesen. Die länglichen Blütenstände, die Kätzchen, sind die männlichen Blüten. In [EE93] wird von einer möglichen geringen Selbstbefruchtungsrate der Sorte Ecker1 berichtet, welche jedoch unter 3 % liegt! Aus persönlicher Erfahrung mit einem isolierten Baum, welcher ein kultivierter Baum ist, liegt bei diesem die Selbstbefruchtungsrate bei etwa 0,5–1 %.
Die Pollen der Edelkastanie sind etwa 16µ m lang – entspricht ungefähr der Größe eines roten Blutkörperchens – sind schwierig zu lagern. Bei warmer und feuchter Lagerung verlieren sie ihre Bestäubungsfähigkeit unter 4 Stunden.
Die Kätzchen kann man bei 20−24 ∘C mit Silica Gel 1-2 Tage trocknen und dann feuchte-dicht verpackt einfrieren [Rut90][Jay74].
In Ländern mit niedriger Luftfeuchte wie Italien gibt es eine effektive Verteilung der Pollen durch Wind und daher Windbestäubung (Anemophilie). In feuchteren Region wie im Westen Frankreichs, werden die Pollen klebrig und es kommt daher laut [Cou12] die Insektenbestäubung, vornehmlich durch Bienen, vor (Entomophilie).
Bienen haben durchaus einen Pollensammelradius von 3 km. Ist es jedoch kalt, wie zur Zeit der Kirschblüte, so wird der Sammelradius von Pollen vielleicht 500 m sein. Bei einem Überangebot an Pollen durch Blütenpflanzen wird der Aktionsradius der Bienen auch gering sein.
Es gibt zwei Arten von Blütenständen: manche bilden nur die länglichen Kätzchen (catkins) aus – diese tragen männlichen Pollen. Bei anderen bilden sich an der Basis des Blütenstandes auch stachelige Blüten aus – das ist der weibliche Teil [Rut90][Lar21].
Abbildung 30: Baum mit beginnender Blüte li, Baum in Blüte re
Abbildung 31: Beginnende Blüte li, männliche und weibl. Blütenstände re
Abbildung 32: Männliche u. weibliche Blütestände im Detail li, Schnitt einer weiblichen Bl. im Detail re
Abbildung 33: Heranwachsender Igel im Detail (2016/KW31) li, Schnitt durch Igel – mit 3 Nüssen re
Die Stamen (Staubblatt-entwicklung) an den Kätzchen ist das Hauptkriterium, ob es sich um eine gute Bestäubersorte handelt:
Astamineae, brahistamineae und mesotamineae Sorten werden als männlich steril eingestuft [Cou12].
Sorten mit lang entwickelten Stamina (longistaminate) sind als Bestäuber einzusetzen. Eine Bestäubersorte sollte im LUV (gegen Hauptwindrichtung bei Pollenflug) männlich steriler Sorten gepflanzt werden. In Plantagen mit männlich sterilen Sorten werden 10 % Bestäubersorten empfohlen.
Gerade die französischen groß-fruchtigen Sorten, die sich auch gut schälen lassen, sind meist Hybriden (Kreuzungen aus C.sat x crenata) und oft männlich steril [Str06]. Das ist z.B. für die Sorte Bouche de Bétizac zutreffend.
Es werden auch sogenannte Duo-Bäume angeboten, auf denen zwei verschiedene Sorten veredelt sind und sich daher selbst befruchten können – wie Doré de Lyon + Belle Epine.
Die männlichen Blüten blühen ca. 7–14 Tage vor den weiblichen Blüten (protandrous). Der Pollenflug dauert normalerweise 3–4 Wochen. Es handelt sich um einen Mechanismus zur Steigerung der Befruchtungseffizienz und zur Risikoverminderung der Blutsverwandtschaft.
|
Etwa 25 % der Blütenpflanzarten in Mitteleuropa sind windbestäubt, der überwiegende Anteil der Blütenpflanzen werden durch Käfer, Falter oder Bienen bestäubt. Bei der Edelkastanie liegt
laut [Cou12] eine Mischform vor, die auch von den klimatischen Bedingungen abhängt: Bei feuchter Luft soll die Insektenbestäubung dominieren. Im Gegensatz dazu wird in [Rut90]
erläutert, dass die männlichen Kätzchen zwar gerne von Insekten aufgesucht werden, die weiblichen aber keine grelle Farbe, keinen Geruch und keinen Nektar aufweisen, damit für Insekten unattraktiv sind und daher die Insektenbestäubung wahrscheinlich eine untergeordnete Rolle spielt. Für eine gute Windbestäubung ist laut [Rut90] ein Baumabstand unter 30 m notwendig. Im gleichen Artikel wird angegeben, dass Bäume ab einem Abstand von etwa 300 m bestäubungstechnisch isoliert sind. Für Windbestäubung ist trockenes Wetter und richtige Windrichtung bei Pollenflug notwendig. Der Bestäuberbaum darf natürlich nicht männlich steril sein! Werden die weiblichen stacheligen Blüten nicht befruchtet, so bildet sich zwar ein Igel aus, dieser enthält aber nur leere Nüsse (siehe Abbildung 34). |
Will man eine Fremdbestäubung mit einfachen Mitteln selbst durchführen, so wartet man solange bis die männlichen Kätzchen am Mutterbaum zu blühen beginnen. Nach weiteren 10 Tagen schneidet blühende männliche Kätzchen von einem Fremdbaum und überstreift die weiblichen Blüten des Mutterbaums mit den abgeschnittenen männlichen Kätzchen. Achtung: Die abgeschnittenen Kätzchen sind ungekühlt – wie schon angeführt – nur wenige Stunden lagerbar [Rut90]!
Die Früchte sind glänzende, dunkelbraune Nüsse. Die Nüsse sind von einem stacheligen Fruchtbecher (Cupula), auch Igel genannt, umgeben. Die Stacheln sind anfangs grün und werden zur Reife gelbbraun. Bei Vollreife öffnet sich der Fruchtbecher mit vier Klappen und entlässt die ein bis drei Früchte/Nüsse (Abbildung 35). Äußerst selten kommen mehr als 3 Nüsse vor. Bei manchen Sorten fällt auch der Fruchtbecher mitsamt den darin enthaltenen Nüssen ab.
Die Wildformen der Edelkastanie sind in kälteren Lagen wie im Wienerwald relativ klein: Abbildung 36 li – Eichenhain: 28 mm, 6 g. Im Burgenland gibt es groß-fruchtige Bäume: Abbildung 36 mi – Rosalia 41 mm, 24 g. Aus Mittelmeerländern wie Italien wird die Edelkastanie exportiert: Abbildung 36 re – Sizilien 43 mm, 32 g.
Der Abschätzung liegen folgende Annahmen zugrunde:
Oberfläche der Baumkrone – Halbkugel mit 10 m Durchmesser: π· d2/2
auf einem m2 Oberfläche 7 Blütengruppen – 7 männliche Blüten pro Gruppe: nBlt/m2= 7 · 7 ≈ 50 / m2
männliche Blüte 10 cm lang: lBlt
50 Staubblätter pro cm männliche Blüte: nStaubb
pro Staubbeutel 5000 Pollen [CM06]: nPoll/Staubb
nPollen = π· d2/2[m2]· nBlt[1/m2]· lBlt[cm]· nStaubb[1/cm]· nPoll/Staubb[1]=
= 3· 102/2· 50· 10· 50· 5000= 3/2· 5· 5· 5· 100.000.000= 3/2· 125· 100 Mio. ≈ 20 Mrd.
Eine Nuss ist ein Samen (eng. Semen). Im Gegensatz zum Apfel (Balgfrucht), wo in der Frucht im Kerngehäuse 5 Kerne bzw. Samen sitzen, oder die Frucht der Feige, welche einige hundert Samen enthält. Aus einer Nuss kann bei natürlicher Vermehrung im Normalfall maximal ein Baum entstehen.
An der Basis der Nuss befindet sich ein weißlicher Fleck, die Trennstelle, an der sich die Nuss von dem Fruchtbecher (Cupula) löst.
Die Bilder 37 wurden in der KW5/2015 gemacht und stammen von am KW40/2014 gesammelten Edelkastanien (Eichenhain).
In der Nuss befindet sich die aus der befruchteten Eizelle (Zygote) gebildete junge Pflanze, der Embryo und zwei Keimblätter (Kotyledonen), die etwa der Hirnoberfläche ähnlich sehen – diese isst man.
Das Gehäuse (Perikarp) der Nuss ist ledrig und gleichmäßig braun oder gestreift. Die Kotyledonen sind von einer gerbstoffhaltigen Haut umgeben, die gute Schälbarkeit dieser Haut ist für die Vermarktung ein wichtiges Kriterium.
Das Hypokotyl beschreibt den unteren Abschnitt einer Pflanze von der Radikula zu den Kotyledonen. Das Epikotyl ist der Abschnitt von den Kotyledonen zum ersten Folgeblatt (siehe auch Abbildung 37 re).
Der Markt oder der Kunde verlangt nach Früchten, die folgende Kriterien maximieren:
große Früchte – weniger als 60 Stück/kg – also ab 17 g pro Nuss
runde oder halbrunde Form
leicht schälbar – die innere, dünne braune Haut der Nuss soll auch maschinell leicht entfernbar sein
gut schmeckend – süß wird in Europa als wohlschmeckend empfunden, bissfest eher nicht, mehlig auch nicht
frühe Reife – frühe Früchte erzielen gute Preise am Markt [Str06]
gleichzeitige Reife – geringerer Ernteaufwand
unverdorben und schädlingsfrei
lange Haltbarkeit – Lagerfähigkeit ohne großen Aufwand
Weizen kann seine Keimfähigkeit bis zu 200 Jahre erhalten (orthodoxer Samen). Der Samen einer Dattelpalme (Phoenix dactylifera) kann über 2000 Jahre keimfähig bleiben (Fundort der Testsamen: Masada-Festung). Im Gegensatz zu unseren Nadelhölzern, haben die Samen der Edelkastanie oder der Eiche diese lange Keimfähigkeit nicht. Sie gehören zu den nur aufwendig lagerbaren Samen („recalcitrante“ Samen) – Eicheln z.B. werden bei −3 ∘C gelagert. Die Nuss der Edelkastanie darf nicht richtig durchfrieren, nicht austrocknen, zu feucht sein oder zu lange gelagert werden um keimfähig zu bleiben. Die Nuss besitzt sogenannte Speicherkotyledonen, welche die Jungpflanze mit Nährstoffen versorgt.
Da die Samen im Herbst reifen, müssen sie in Mitteleuropa überwintern und sind daher zumindest für ein halbes Jahr Keimfähigkeit ausgelegt – was auch der normalen Lagerzeit der Nüsse/Samen entspricht. Laut [Jay74] kann man die Nüsse bei optimalen Bedingungen auch bis zu 3 1/2 Jahre lebensfähig erhalten und eine Einlagerung von 1 1/2 Jahren erscheint praktikabel. Aus eigener Erfahrung sind die Nüsse nach einer Lagerung von 11 Monaten und bei etwa 5 ∘C schon stark angetrieben.
Für die Lagertemperatur wird ein Bereich von −3 ∘C bis +4,5 ∘C angegeben [Rut90]. Möglichst niedrige Temperaturen verlangsamen die Entwicklung von Schimmelpilzen und verzögern die Entwicklung der Samen. Andererseits wird vermutlich auch die Keimungsrate sinken.
Wobei letztendlich +3 – +4∘C laut [Gos07] [dIFdL14] als Richtwert betrachtet wird. Bei der Lagerung sollte ein Gasaustausch möglich sein, also keine luftdichten Behälter verwenden. Die Luftfeuchtigkeit im Behälter sollte bei etwa 40 % liegen [oA08].
Genauer betrachtet ist die Kältetoleranz der Nüsse von Art zu Art verschiedenen und innerhalb einer Art sind Nüsse aus kälteren klimatischen Zonen auch kälteresistenter. In [SSH+12] wird beschrieben, dass die Kälteresistenz bei einer definierten Zellmortalitätsrate von 50 % für die Castanea dentata in klimatisch Wärmeren Gebieten wie Kentucky bei etwa −12 ∘C und in klimatisch kälteren Gebieten wie Maine bei etwa −16 ∘C liegt (Tm). Für die Castanea mollissima wird die Kälteresistenz mit etwa −6 ∘C angegeben. Die angegebenen Resistenzwerte Tm basieren nicht auf Anzuchtversuche nach einer definierten Kältebehandlung, sondern es wurde über Leitfähigkeitsmessungen von kurzzeitig kältebehandelten Nusstückchen die Resistenztemperatur Tm indirekt bestimmt.
Es wird zusätzlicher Elektrolyt, hauptsächlich Kaliumionen K+, aus dem Zellinneren durch Dehydrierung und geplatzten Zellwänden freigesetzt und damit die Leitfähigkeit der Probe erhöht. Im Prinzip wird für jeden Kältepunkt nach der Kältebehandlung eine Leitfähigkeitsmessung mit den Zellproben durchgeführt und nach Zerstörung der restlichen Zellwände (z.B. durch Aufkochen) eine zweite Leitfähigkeitsmessung durchgeführt. Die Kurve mit der Differenzleitfähigkeit über der Temperatur dient als Basis für die Berechnung der Kälteresistenz Tm (REL – Relative Electrolyte Leakage).
Vor der Kältebehandlung wird kein spezieller Elektrolyt in die Zellen eingelagert oder spezifische Kältezyklen gefahren wie es bei einer Kyrokonservierung von Pflanzenmaterial erfolgt.
| Edelkastanien sind keine Frostkeimer! Die Edelkastanie braucht keinen Frost um zu keimen und ist auch kontraproduktiv, da manche Nüsse auch schon bei +4 ∘C leicht austreiben. In diesem Stadium sind die Nüsse extrem frostempfindlich. Lässt man die Nuss richtig durchfrieren, so ist sie nicht mehr keimfähig – im Gegensatz zur Rosskastanie, welche ein Frostkeimer ist. Als Richtwert möchte ich aus eigener Erfahrung hier eine minimale kurzzeitige Kälteresistenz (eine Nacht) von mindestens −8 ∘C für Castanea sativa Nüsse und deren crenata Einkreuzungen angeben. |
Die Kastanie kann man auch direkt nach der Ernte zum Keimen bringen. Es dauert aus eigener Erfahrung bei Raumtemperatur so ca. 4 bis 8 Wochen bis zur Keimung. Die Ankeimrate ist jedoch höher, wenn man die Kastanien z.B. im Kühlschrank bei +5∘C min. 6–8 Wochen lange stratifiziert.
Die oft angegebene Dormanz der Edelkastanie dürfte aber eher eine unterordnete Rolle spielen. Auffällig und auch bekannt ist, dass nach einer gekühlten Lagerung der Nüsse, diese innerhalb von 1–2 Wochen gemeinsam keimen.
Die kalte Lagerung wird auch verwendet, um die Keimung hinauszögern, damit man die Samen direkt im Frühjahr ins Freie auszusetzen kann. Setzt man die Nüsse im Herbst direkt ins Freie in die Erde, so wird die Ausbeute stark sinken, da die notwendigen Rahmenbedingungen wie Bodentemperatur, Feuchtigkeit (bei zu viel Nässe werden sie verfaulen) oft nicht passend sind, und außerdem die Nüsse in der nahrungsarmen Zeit gerne von Tieren aus den Boden ausgegraben und gefressen werden.
Abbildung 39: Aussortieren li, Lagerung re
Man sollte die gesammelten Nüsse möglichst innerhalb von 3 Tagen richtig lagern oder einpflanzen.
Die Nüsse werden über den Winter bei maximal ( 3−5 ∘C) im Kühlschrack oder Kühlkammer gelagert. Zunächst werden die gesammelten Nüsse aussortiert: alle schwimmfähigen (Abbildung 39) Kastanien werden verworfen. Danach trocknet man die Kastanien auf einem Tuch. Die Edelkastanien können auch mit einem Nagellack farbig markiert und einer Zuordnungsliste verwaltet werden.
Einstiegsvariante: Man packt die Eedelkastanien, gemischt mit etwas Hochmoortorf, in Zipper-Säcke. In einem 3 l Zipper nicht mehr als einen halben kg Nüsse. Als Obergrenze nicht mehr als 1 kg in einem Paket, da sonst die erzeugte Wärme zu langsam abgeführt werden kann. Man verschließt die Säcke nur zu drei viertel. Außerdem legt man ein angefeuchtetes Blatt einer Küchenrolle bei.
Man beschriftet die Packung mit dem Funddatum, Fundort etc., versucht die Kastanien flach auszurichten und legt das Paket in die Gemüselade des Kühlschranks. Man sollte sich dabei im Klaren sein, dass sich Larven des Edelkastanienbohrers auch durch Plastiksäcke fressen können...
Variante mit Box: Diese Variante hat den großen Vorteil, dass die Nüsse ausgerichtet eingeschlichtet werden. Und die Pfahlwurzelnbei der Keimung ungehindert wachsen können, was wichtig bei einer Ammenveredelung ist . In eine Plastikbox z. B. mit l x b x h = 34 x 19,5 x 11,5 cm werden bis zu 30 Nüsse – je nach Größe, einlagig eingeschlichtet.
Hinweis zur Feuchtigkeit der Box: Zunächst eine angfeuchtete Schicht Torf einbringen, dann eine trockene Schicht aufbringen – Schichtung jeweils etwa 6 und 2 cm hoch. Die obere trockene Schicht mit einer Blumenspritze anfeuchten, die Edelkastanien systematisch auflegen, etwas andrücken und wieder eine trockene Torfschicht aufbringen. Die Nüsse sind nicht mehr sichtbar. Danach den Deckel aufdrücken und die beschriftete Box im Kühlschrank lagern.
Wichtig für den ersten Versuch: meist werden die Kastanien zu feucht eingelagert – der Torf soll noch bröselig sein. Werden die Nüsse zu feucht gelagert, so werden sie trotz des Torfes zu schimmeln beginnen!
Alle 2–4 Wochen sollte man den Sack/Box kontrollieren und eventuell angeschimmelte Nüsse entfernen.
Mit einer Blumenspritze etwas Feuchtigkeit hinzufügen. Bei der Sackmethode diesen nicht wenden. Die Ausrichtung der Nüsse sollte beibehalten werden! Nach etwa 3–4 Wochen wird bei den ersten Nüssen die Radikula (vielleicht bei 3 % der Nüsse) sichtbar. Nach 3 Monaten ( 5 ∘C) werden bei ca. 50 % der Nüsse Radikeln sichtbar sein.
Abbildung 40: Vorbereitung der Low-Tech Variante eines Pflanztopfes: Drainagelöcher li, Pflanzerde mit aufgelegtem Keimling re
Abbildung 41: angekeimte Nüsse: mit kurzer Radikula li, Radikula mit kleinen Seitenwurzeln re
Die angekeimten Nüsse kann man herausnehmen und in Anzuchtsubstrat, siehe Abschnitt 5.3 gezeigt oder auch zunächst in reinen Torf setzen. Als Einstiegsvariante kann man aufgeschnittene Tetra-Packungen mit ein paar Löchern am Boden einsetzen (Abbildung 40 li). Professioneller ist es, mehrere angetriebene Nüsse in Kisten mit mindestens 15 cm zu setzen.
Nach ca. 6–8 Wochen kann man, wenn man will, alle kühl gelagerten Nüsse – auch die ungekeimten, einpflanzen. Die Kastanien werden im Topf/Kiste flach aufgelegt und ca. 0,5 cm mit Substrat locker bedeckt. Die Keimung wird bei Raumtemperatur in 1–2 Wochen erfolgen. Insgesamt sollte man auf eine Keimungsausbeute von mehr als 80 % kommen.
Spätestens, wenn die Bäumchen im Frühjahr 20–40 cm erreicht haben, gehören diese vereinzelt oder aus den Tetra-packungen entfernt und wie in Abschnitt 7 beschrieben eingetopft.
Sind die Edelkastanien vor dem Aufsammeln schon längere Zeit am Boden gelegen und das Klima trocken oder frostig, so kann die Keimrate massiv sinken.
Solange nur die Radikula (Abbildung 41 li) vorhanden ist, kann man die angekeimten Nüsse problemlos umsetzen. Wenn Ansätze von Seitenwurzeln (Abbildung 41 re) vorhanden sind, sollte man die Kastanien nicht mehr wurzelnackt umbetten.
Die Abbildung 42 zeigt Wildformen – bei Gloggnitz (gesammelt KW44/2014, gepflanzt: KW2/2015), Eichenhain (KW40/2014, KW52/2014) und Rosalia (KW40/2014 / ca. KW44/2014) – Aufnahme von KW6/2015.
Autovegetativ werden Pflanzen z.B. über Steckhölzer oder Abrisse vermehrt. Die gesamte neue Pflanze ist genetisch ident. Heterovegetativ werden normalerweise Pflanzen durch Methoden wie Okulation oder Kopulation vermehrt – die neue Pflanze besteht genetisch aus verschiedenen Pflanzenbestandteilen, die zusammengewachsen sind. Normalerweise ist bei einem veredeltem Baum die Unterlage (Wurzelstock und ein Teil des Stammes) genetisch verschieden zur Krone. Das Verbinden genetisch identer Pflanzteile macht nur in Ausnahmefällen wie zur Rettung eines verletzten Baumes ([Gau91]) Sinn.
Generell sind Buchengewächse wie die Edelkastanie schwieriger zu veredeln als Rosengewächse wie Apfel und Birne.
Details über historische und noch heute verwendete Veredelungsmethoden findet man in [Gau91] mit:
Ablaktieren – durch Anplatten, in Vertiefung, mit Gegenzunge
Propfen – in Seite mit Geißfuß, zw. Rinde und Kambium, seitlich in Spalt, in gespaltene Endknospe, Gipfeltriebe, in unbewurzelte Stecklinge
Kopulieren – einfach, mit Gegenzungen, durch Schäften, Anplatten
Okulation – durch T-Schnitt, mit verkehrtem T-Schnitt, durch Anplatten, durch Röhrlen.
Als ebenfalls nicht mehr unter Copyright stehendes Buch über Anbau inklusive Veredelungsmethoden ist auch [Bai20] zu empfehlen.
Einen gut bebilderten Überblick der Veredelungsmethoden der Edelkastanie findet man in [Kö15b], [JFS12] und unter [Viv14] eine Playliste von Videos über Pflege und Veredelung.
Veredelungen wurden schon in der Antike durchgeführt. So wird in der Bibel in dem Römerbrief von Paulus: Röm 11:17-24 vom wilden und vom edlen Ölbaum gesprochen. Das Wort einsetzen/impfen (ἐγκεντρίζω) wird 6 mal verwendet.
|
Bei allen Veredelungsmethoden muss das Kambium beider Pflanzteile zur Deckung/Teildeckung gebracht werden um ein Verwachsen zu ermöglichen.
Veredelungen sollten immer bei moderater Temperatur und nie bei direkter Sonneneinstrahlung durchgeführt werden – also am Morgen, im Schatten oder
in einem kühlen Raum. Regenperioden sind zu vermeiden, da die Feuchtigkeit der Erde/Substrat gering sein sollte, um keinen massiven Saftstrom in der
Unterlage anzuregen [MRW12]. Die Schnittstellen sollten so schnell wie möglich zusammenfügt werden. Beim Fixieren des Edelreises auf die Unterlage muss darauf geachtet werden, dass es nicht zum Einschnüren und damit zum Zusammenbruch von Gewebeteilen – insbesonders von Leitgewebe wie das Phloem – kommt. Die Schnittwerkzeuge sind mit Isopropanol oder einem anderen Desinfektionsmittel zu desinfizieren. Außerdem sollte man die Schnittstellen nie berühren. |
Während für die Forstwirtschaft normalerweise generativ vermehrte Pflanzen verwendet werden, sind im Erwerbsanbau faktisch nur vegetativ vermehrte Edelkastanienbäume im Einsatz. Wenn Sämlinge verwendet werden, bilden sie nur den Wurzelstock und einen Teil des Stammes.
Bei vegetativ vermehrten Edelkastanien kann auf eine Veredelung verzichtet werden, wenn keine spezielle Unterlage (Wurzelstock) gebraucht wird. Wurzelstöcke, welche gegen Tintenkrankheit immun sind, werden oft für Bäume verwendet, die diese Eigenschaft nicht aufweisen (siehe auch Abbildung 43).
Vermehrung aus Steckhölzern funktioniert mit brauchbarer Ausbeute nur bei Jungbäumen als Spender (siehe auch Abschnitt 9.8.2).
Steckt man einen Weidenast in feuchte Erde, so werden sich innerhalb einiger Tage Wurzeln ausbilden. Die Weide produziert das Phytohormon IBA 4-(Indol-3-yl)buttersäure (C12H13NO2) und Salicin in größeren Mengen selbst.
Salicin wird bei Einnahme in den Körper zu Salicylsäure (C7H6O3) umgebaut. Salicin ist für die pflanzliche Abwehr von Pathogenen verantwortlich und hat auch eine antimikrobielle Wirkung. Salicylsäure ist als Konservierungsmittel in Lebensmitteln verboten. IBA ist ein Phytohormon aus der Gruppe der Auxine, welches das Längenwachstum und die Wurzelbildung anregt (siehe auch Abschnitt 15).
Stellt man abgeschnittene Triebe von der Edelkastanie im Winter ins Wasser, so werden eventuell Blätter und auch Blütenansätze gebildet (Abbildung 44) und es kommt vielleicht auch zur Kallusbildung, jedoch normalerweise zu keiner Wurzelbildung.
Auxinprotektoren verhindern die einfache und rasche Wurzelbildung der Edelkastanie. In [Ost08] wird über eine erfolgreiche Stecklings-Vermehrung der Esskastanie etwas detaillierter berichtet. Die 12 cm langen Stecklinge wurden in ein Torf- Sandgemisch gesteckt. Vorbehandelt wurden die Stecklinge mit einem Bewurzelungshormon und Zusätzen. Als Bewurzelungshormon wurde 0,5 % IBA+Euparen auf Talkumbasis verwendet. Die Pflanzung Ende Mai ergab die beste Ausbeute.
Erklärungen: Euparen (Bayer, CropScience) enthält den Wirkstoff Tolylfluanid und ist ein Fungizid. Die Stecklingsvermehrung von 3–4 jährigen Bäumen (green cuttings) hat aber an Bedeutung für die Vermehrung von Hybridsorten gewonnen [Cou12]. Eine gute Zusammenstellung über Techniken der Stecklingsvermehrung findet man in [HK14]
Abbildung 44: Steckling im Wasser: mit Kallusbildung li und Bildung von Blütenständen re
Im Abschnitt 15 werden verschiedene Bewurzelungspräparate für Stecklinge angeführt und deren Handhabung angeführt.
Diese Methode, eine Autovegetative Methode, generiert genetisch idente Abbilder der Mutterpflanze. Sie wird im Erwerbsanbau für die Produktion von Veredelungsunterlagen (vor allem tintenkrankheitsresistente Unterlagen) für spätere Veredelung wie Okulation, Chip-Budding, Kopulation eingesetzt.
Abrisse werden im Prinzip folgendermaßen hergestellt: man kappt eine Edelkastanie im Herbst knapp oberhalb der Erde. Die sogenannte Mutterpflanze wird im Frühjahr mehrere Jungtriebe bilden, auf jedem dieser Triebe wird Ende Mai bis Juni ein metallischer Ring am unteren Ende aus Draht um den Trieb herumgebogen. Sinn: wächst der Trieb, so wird der Trieb durch den Ring eingeschnürt, wobei die Wasserversorgung von der Mutterpflanze noch funktioniert, aber die Phytohormone und Kohlehydrate von den Triebspitzen nicht mehr zu den Wurzeln der Mutterpflanze über das Phloem abfließen können. Dieser Stau und zusätzlich über den Ring aufgestrichene Auxine bewirken ein Wurzelwachstum oberhalb des Einschnürringes (Girdle).
Als Bewurzelungshilfe wird ein Auxin wie IBA mit einer Trägersubstanz wie Vaseline (etwa 2–3 g/kg) eingesetzt (siehe auch Bewurzelungsgel im Abschnitt 15.2).
Als letzter Schritt wird der ganze Wurzelstock mit Erde angehäufelt – also fast mit Erde bedeckt. Die Jungtriebe werden weiterwachsen und außerdem oberhalb der Ringe Wurzeln gebildet. Diese Jungtriebe werden im Herbst von der Mutterpflanze abgerissen bzw. abgeschnitten, wobei die Ringe auch eine Sollbruchstelle bilden.
Diese Methode wird immer erfolgreicher bei der Vermehrung von Edelkastanien in Baumschulen in Südeuropa eingesetzt [Viv12] [Viv13] [dIFdL14]. Aus Sämlingen gezogene Unterlagen werden immer mehr verdrängt.
Im Obstbau wird über die Unterlage die Größe des Baumes definiert. Beim Apfel werden schwach wachsende vegetativ vermehrte Unterlagen wie M9 (2,5–4,5 m) und im Erwerbsobstbau für Säulenbäume z.B. die Unterlage M26 verwendet. Stark wachsende Unterlagen werden aus Sämlingen wie Antonowka, Bittenfelder oder Grahams gezogen und für große Hochstammbäume verwendet.
Der veredelte Teil des Baum entwickelt sich aus einer oder mehreren Knospen (Edelaugen). Die geschnittenen veredelten Triebe (Edelreiser) sollen für die meisten Veredelungsmethoden mindestens bleistiftstärke mit 8 mm und eine Länge von mindestens 30 cm und einige Edelaugen aufweisen. Die Triebe sind sorgfältig auszuwählen und nicht am Baum wild herumzuschneiden!
Allzu dünne Edelreiser haben zu wenig Reservestoffe wie Kohlehydrate eingelagert. Von gespeicherten Kohlehydraten hängt unter anderem auch die Intensität der Kallusbildung ab.
Die besten Knospen befinden sich meist in der Mitte des Edelreises. Igel-tragende Reiser sind weniger geeignet, da sich am Ort der Fruchtbildung am Reis keine schlafende Blattknospe bildet. Igeltragende Reiser zeigen im Winter an der Rinde Narben der abgefallenen Fruchtstände (Abbildung 44 re). Die vegetativen Knospen am Reis sind flacher. Die Reiser müssen elastisch und gesund sein. Triebe unter der Veredelungsstelle sind als Edelreis natürlich dafür ungeeignet. Ältere Bäume haben leider nicht so lange und schlanke Jahrestriebe und sind daher dann oft ein Kompromiss.
Ein– bis Zweijährige Triebe, von zurückgeschnittenen Bäumen sind ideal. Die weichen Wassertriebe sollte man nicht verwenden.
Und eine gute Affinität zur Unterlage haben – gemeint ist hier die Verträglichkeit zwischen dem Edelreis und der Unterlage.
Edelreiser für Kopulation werden in Mitteleurope meist Mitte Dezember bis Ende Februar an einem frostfreien Tag geschnitten. Meist ist Ende Jänner die beste Zeit für den Schnitt. Jedenfalls darf der Saftstrom darf noch nicht begonnen haben. Die Reiserstärke muss bei der Einfachen Kopulation der Unterlage gleich stark sein.
Edelreiser für Okulation im Sommer werden meist unmittelbar vor der Veredelung geschnitten. Wird eine Okulation im August durchgeführt, so sind schlafende Augen zu verwenden, welche erst im nächsten Frühjahr austreiben.
Edelreiser für die Ammenveredelung/Ammenvermehrung werden auch Mitte Dezember bis Ende Februar in einer Stärke von meist 4–6 mm geschnitten. Siehe auch Abbildung 58.
Die geschnittenen Reiser werden beschriftet, gebündelt und in feuchtes Papier, Moos, Torf oder Sand an der eingeschlagen und in Plastiksäcke verpackt und bei 1 − 4 ∘C im Dunkeln bis zum Veredelungstermin gelagert. Die Reiser dürfen weder austrocknen noch zu nass liegen. Auch sind die Reiser von Ethylen bildendem Obst und Gemüse wie Äpfel, Birnen, Bananen, Paradeiser getrennt zu lagern, da Ethylen (Äthylen, Ethen) die Knospen schädigt. Ethylen ist ein Phytohormon und bewirkt eine Hemmung des Längenwachstums, die Fruchtreifung und den Abwurf der Blätter im Herbst.
Diese Veredelungsmethode auch einfache Kopulation genannt, wird am Ende Ruhezeit der Pflanze angewandt (Spätwinter oder Frühjahr). Die Methode wird frühestens angewandt, wenn die Knospen der Unterlage zu Schwellen beginnen. Bei den Edelreisern müssen die Knospen noch ruhend bzw. geschlossen sein. Das zurechtgeschnittene Edelreis sollte sicherheitshalber mindestens 2 Knospen aufweisen. Mehr als vier Augen sind ungünstig, da dann die Versorgung des Edelreises während der Verwachsungsphase meist unzureichend ist.
Eine am Edelreis und auch an der Schnittstelle der Unterlage gegenüberliegenden Knospe soll das Anwachsen fördern.
Das Edelreis und die Unterlage sollten annähernd gleiche Durchmesser haben und beide werden schräg (etwa 4–5 so lang wie Durchmesser) abgeschnitten und dann so miteinander verbunden, dass die Kambium-Schichten aufeinander liegen (Abbildung 46). Als Stammstärke für eine normale Schrägschnittkopulation wird bleistiftstärke bis besenstielstärke (etwa 8 bis 25 mm in Durchmesser) an der Veredelungsstelle angegeben. Die beiden Teile wurden früher mit Bast, Gummiband formschlüssig verbunden und mit Wachs, Wundbalsam steril abgedichtet, Heute wird im professionellen Umfeld meist mit einem Veredelungsband wie Buddytape von Aglis, Medifilm, Parafilm Grafting Tape, gearbeitet, da eine sterile Abdichtung gleich über das dehnbaren Band erfolgt. Bei dünneren Edelreisen und niedrigen Temperaturen ist auch Parafilm M (Abbildung 47) eine gute Wahl.
Treibt das Edelauge aus, so wird der Stummel zunächst als Führung zum Aufschulen des Jungtriebes verwendet. Der Stummel wird nach einigen Wochen schräg oberhalb des veredeltem Jungtriebes abgeschnitten und die Wunde mit Baumwachs, Parafilm etc. verschlossen und der Trieb an einem Stützstab weiter aufgeschult (Abbildung 47). Zusätzliche Triebe werden entfernt.
| Es ist abzuraten ein langes Edelreis mit terminalen Knospen auf die Unterlage zu kopulieren, da die Veredelungsstelle in der Verwachsungsphase das Edelreis meist nicht ausreichend versorgen kann. Außerdem befinden sich am terminalen Ende meist Blütenknospen, die ein Längenwachstum des veredelten Baumes verhindern. |
Bei dieser Methode wird ein Auge (Knospe) des Edelreises mit der Unterlage verbunden, daher auch Veredeln mit einem Auge genannt.
Die Unterlage muss im Saft sein. Die Rinde (Borke + Bast) muss sich einfach (vom Kambium) lösen, was im Frühjahr, Spätsommer und Herbst (etwa April bis September) der Fall ist. Die Methode wird bei Jungbäumen mit glatter Rinde angewandt. Bei der Frühjahrsokulation werden die gekühlt gelagerten Edelreiser des letzten Jahres verwendet. Bei der Sommerokulation verwendet man schlafende Knospen (kleine Knospen oberhalb eines Blattstieles sitzend), welche erst nächstes Jahr austreiben. Die Unterlage muss jedenfalls noch ein gutes Monat im Wachstum sein, um ein gutes Verwachsen zu gewährleisten. Aber die Veredelung ist schon so spät durchzuführen, dass die Edelaugen nicht mehr austreiben. Das Edelreis sollte etwas dünner oder gleich stark wie die Unterlage sein.
Im Erwerbsanbau wird bei dieser Methode, hier exemplarisch der gesamte Produktionsprozess nach [EE93] (1993) beschrieben, folgendermaßen vorgegangen: Die Nüsse werden im Herbst im Torf eingelagert und im Frühjahr die angekeimten Nüsse ins Freie ausgesetzt. Diese wachsen über die Sommersaison und im Herbst werden die ca. 50 cm großen Pflanzen entblättert, ausgegraben und auf ca. 30 cm zurückgeschnitten und über den Winter kühl gelagert.
Im darauffolgenden Frühjahr werden die rückgeschnittenen Bäume erneut ausgepflanzt und im August ab einer Stammstärke von ungefähr 10 mm okuliert.
Zur Okulation wird das Auge flach mit einer Länge von ca. 2 cm aus dem Edelreis geschnitten. Das im Auge verbleibende Holzschildchen, soll beim Herauslösen brechen und nicht das Augenkernholz herausreißen. Die Schnittstellen nicht angreifen und das Okuliermesser desinfizieren!
An der Unterlage wird knapp über dem Boden (5-10 cm) ein ca. 2 cm großes T geschnitten und das Auge eingesetzt. Der nach oben überstehende Teil des Auges wird am T-Dach abgeschnitten. Danach wird die Okulationsstelle mit einem durchsichtigem Veredelungsband wie z.B. Buddytape, Parafilm M oder Medifilm verbunden, sodass das Auge fest anliegt und nicht austrocknet. Ab dem neunten Tag sollte das Auge schon gut auf der Unterlage haften und Kallus-Wachstum sichtbar sein. Das Auge wird hoffentlich im Frühjahr antreiben und als neuer Trieb seitlich nach oben wachsen, wobei der Jungtrieb zu den Stammstummel hin gebunden werden kann. Nach einigen Wochen wird Der Stammstummel beim Auge, schräg vom Jungtrieb nach unten weg abgeschnitten und die Schnittstelle mit Wundbalsam versorgt.
Nach 2 Sommerperioden hat der okulierte Baum ca. 1,5–2 m erreicht und ist im Herbst verkaufsfertig. Insgesamt dauert es vom Einlagern der Nüsse bis zum verkaufsfertigen Baum etwa 3 Jahre. Das Wachstum der Pflanzen erfolgt im Freien auf dem Feld.
Detaillierte Ausführungen zur Methode findet man in der Diplomarbeit [EE93]. Die Aufbereitung des Edelauges wird ausführlich bebildert in [Kö15b] gezeigt.
Für diese Veredelungsmethode, welche eine Form der Okulation ist, wird auch Plattenokulation oder Chip-Veredelung genannt, wird ein Span mit Knospe des Edelreises mit der Unterlage verbunden.
Im Erwerbsanbau wird bei dieser Methode nach [Kö15b] folgendermaßen vorgegangen: Die Nüsse werden im Herbst in Torf vereinzelt eingelagert und auch gemeinsam vorgezogen. Im Frühjahr werden die kleinen Pflanzen schon große Töpfe umgesetzt. Mitte August wird durch Chip-Budding die etwa 50 cm großen Pflanze mit einem Edelauge versehen. Der eingesetzte Span wird mit einem durchsichtigen Veredelungsband wie z.B. Buddytape, Parafilm M oder Medifilm verschlossen. Nach etwa 5 Wochen wird das Band entfernt. Im Spätherbst werden die Pflanzen etwa 10 cm über dem Edelauge abgeschnitten und zum Überwintern mit den Töpfen in große Kisten verpackt. Die Apikaldominanz würde sonst ein Austreiben eines Seitentriebes am Edelauge verhindern. Im nächsten Frühjahr wird die wird die Pflanze am Edelauge austreiben, der veredelte Jungtrieb zunächst am Reststummel aufgeschult und der Stummel später abgeschnitten. Sollte im Frühjahr kein Austrieb am Chip erfolgen, so kann im Sommer oder auch schon im Frühjahr die Veredelung nochmals erfolgen. Im Herbst sollte der veredelte Baum ca. 1,5–2 m erreicht haben und damit verkaufsfertig [Kö15b] sein. Vom Einlagern der Nüsse bis zum verkaufsfertigen Baum dauert es bei [Kö15b] (2015) im Normalfall 2 Jahre.
Bei der Cadillac-Methode wird das untere Ende des Edelreises keilförmig angespitzt und in einen vorgeschnittenen Spalt nahe am Wurzelhals in die Unterlage hineingedrückt. Diese Veredelungsmethode wird vor allem bei Walnussbäumen am Tisch angewendet. Sie wird aber auch in Frankreich zur Veredelung von Edelkastanienbäumen angewendet. Joseph Capus entwickelte die Cadillac-Methode in landwirtschaftlichen Forschungsanstalt der Gemeinde Cadillac in der Gebietskörperschaft Gironde.
Die Ammenveredelung (eng. Nurse-Seed Grafting) ist eine vegetative Vermehrungsart, bei der die Nährstoffe des Samens genutzt werden um das Edelreis mit Nährstoffen zu versorgen. Die Vermehrung über einen Ammensamen ist oft eine alternative Vermehrungsart für Pflanzen, welche schwer durch Steckhölzer zu vermehren sind und große Samen haben [Lew75] [Sch10].
Man versucht genauso wie beim Kopulieren, eine möglichst gute Verbindung zwischen Unterlage, in diesem Fall die angekeimte Nuss, und dem Edelreis zu erzeugen und damit über Kallus-Bildung ein Verwachsen der Übergangsstelle zu ermöglichen. Entwickelt das Edelreis eigene Wurzeln, so spielt die Unterlage (Samen/Nuss) in diesem Fall keine oder geringe Rolle im weiteren Leben der Pflanze. Das heißt auch, man braucht dann nicht auf Kompatibilität zwischen Unterlage und Edelreis zu achten. Das ist leider bei Edelkastanie nicht der Fall, Wurzeln bilden sich leider nur von der Nuss ausgehend aus.
Das Edelreis wird zunächst über eine Amme versorgt, aber Teile der Amme, die Wurzeln sind bleibend sind und damit ist es streng genommen keine klassische Vermehrung über eine Amme. Was Jaynes und Messner auch richtigstellen wollten und sie schlugen daher 1967 als Bezeichnung eng. Nut Grafting anstelle als Ersatz von eng. Nurse Seed Grafting für die Edelkastanie vor [Vie74]. Der Vorschlag wurde aber nie richtig angenommen.
Insgesamt werden in der Literatur vier Arten beschrieben: das Edelreis wird direkt in die angekeimte Nuss (in den Keimling), in die Spitze der Radikula (engl. inverted radicle method), ins Hypokotyl oder ins Epikotyl der angekeimten Nuss platziert.
Ammenveredelung wird z.B. auch bei Avocados und Kamelien angewandt.
Die Vorbereitung der Edelreisstifte sind für alle vier Methoden in etwa gleich. Als Werkzeug, Hilfsmaterial und Pflanzsubtstrat werden benötigt: kleine Schere, Messer, Isopropanol-Desinfektionstupfer aus der Apotheke gehen auch, Parafilm – kein Isolierband verwenden, als Gewächshaus durchsichtige Plastiksäcke mit Zip ab 3 l oder größere Plastikboxen, Hochmoortorf oder ein Hochmoortorf-Vermiculit-Gemisch.
Pflanzliches Material: größere, angekeimte Nüsse, wobei die Radikula mindestens 4 mm dick, mindestens 3 cm lang und noch keine Seitenwurzeln haben soll. Die Edelreiser dazu passend, in etwa der gleichen Stärke.
Die Edelreiser, Triebe des letzten Jahres, werden in der Winterperiode, meist Ende Jänner, vom Baum geschnitten und im leicht feuchten Torf und in Plastiksäcke gekühlt gelagert (genaueres dazu siehe Abschnitt 9.8.4). Oder man schneidet die Edelreiser im Winter unmittelbar vorher – geht bis zum Anschwellen der Knospen, also manchmal bis in den April hinein (zur Ausbeute siehe auch Abschnitt 9.8.9).
Die Schneidwerkzeuge bei Baumwechsel desinfizieren! Das Edelreis muss elastisch und die Knospen gefüllt sein.
Es sollte versucht werden möglichst steril zu arbeiten, so kann man das Schneidewerkzeug mit Isopropanol desinfizieren. Die Schnittstellen von Nuss und Reis natürlich nicht berühren!
Die in der Box oder Zipper angekeimten Nüsse etwa eine Stunde oder etwas länger auf Raumtemperatur aufwärmen lassen. Dann die Nüsse vorsichtig aus dem Torf herausziehen oder besser die Box vorsichtig ausleeren und die Torfreste abspülen. Die gereinigten Nüsse mit einer Küchenrolle abtupfen und in eine feucht gehaltene Box legen.
Abbildung 60: vorbereitete angekeimte Nüsse li, Werkzeug re
Das Zuschneiden der Edelreisstifte kann man bei allen drei Methoden gleichartig durchführen. Die Stifte nicht auf Vorrat schneiden, sonden das Anspitzen unmittelbar vor dem Verbinden machen. Es werden etwa 5–7 cm langen Stifte mit je einer vegetativen Knospe aus dem Edelreis geschnitten, je nach Knospenverlauf etwas länger oder kürzer. Bei einer Reiserstärke von etwas 4 bis 6 mm, sollte das keilförmige Ende nicht länger als 1 cm sein. Wie man in Abbildung 61 sieht, ist die Schnittführung nicht geradlinig, sondern leicht konkav.
Abbildung 61: Ammenveredelung: richtig geschnittene Edelreisstifte
Für die vielen Versuche habt mich die Familie Klug, die Familie Pucher und die Familie Laimer mit Edelreiser von verschiedensten Sorten versorgt. Das ermöglichte eine kleine Versuchsreihe mit Reisern von bekannten Sorten wie Bouche de Bétizac, Colossal, Dore de Lion, Ecker1, Ecker2, Bournette, H2, Hacker, Marrone di Segni, Südtiroler Gelbe auf Nüsse von Wildlinge und Verkaufnüssen. Vorab, die Reiser von den Sorten Bouche de Betizac und die Befruchtersorte H2 haben die besten Anwuchsleistungen.
Danke für die Unterstützung!
Der günstige Veredelungzeitbereich bzw. Anzucht mit Amme liegt für Mitteleuropa von KW12 bis KW15. Die maximale Außentemperatur sollte im Schatten schon bei 20 ∘C liegen.
Als erster Versuch der Vegetativen Vermehrung durch Amme wurde die Methode direkt in den Keimling durchgeführt (engl. auch Nut Grafting). Die folgenden Arbeitsschritte zeigen detailiert auf, was man alles beachten sollte.
Diese Methode wurde von Moore 1963 beschrieben.
Von der angetriebenen Nuss sind von der Spitze ausgehend ca. 2 mm abzuschneiden. Mit einem Messer wird dann etwa 1 – 1,5 cm senkrecht kreuzweise in die Schnittstelle gestochen. Die gesamte Nuss darf dabei nicht gespalten werden! Das keilförmige Ende des Edelreises wird in die vorbereite Schnittstelle eingedrückt (siehe auch Abbildung 9.8.9) und die Schnittstelle mit Parafilm maximal zweimal umwickelt.

Abbildung 62: Ammenvermehrung: direkt in die Nuss
Die Methode ist meiner Meinung nach nur für Grobmotoriker sinnvoll. Ich habe sie nicht weiter verfolgt, obwohl die Verbindung zum Edelreis gut war, da die Ausbildung des Wurzelstockes stark gestört wird–so kann sich keine Pfahlwurzel ausbilden
Das engl. Inverted Radicle Grafting wird von Park 1968 beschrieben. Dazu wird die Spitze der Radikula abgeschnitten und die Radikula in der notwendigen Länge längs gespalten. Danach wird das keilförmige Ende des Edelreises hineingelegt und mit Parafilm straff umwickelt und damit verschlossen (engl: Inverted Radicle Grafting) ). Nicht vorarbeiten, immer nur eine Nuss und ein Edelreistückchen vorbereiten und alle Schnittbereiche zügig luftdicht verschließen.
Dazu von der Parafilmrolle etwa einen 7–10 mm breiten Streifen herunterschneiden. Am oberen Ende zum Edelreis zu wickeln beginnen und oben möglichst nur eine oder zwei Lagen aufbringen, sodass die Wurzeln auch durch den Parafilm wachsen können!
In der Mitte des Einschnittes hin so fest wickeln, dass der Parafilm fast vor dem Abreißen ist. Den Vorgang des Umwickelns vielleicht vorab an einem Aststückchen üben.
Vorteile:
Nachteile:
Abbildung 64: Ammenvermehrung: invertierte Radicula: Nuss u. Edelreis li, Verbindungsstelle re
Diese Methode habe ich nicht weiter verfolgt, weil die verwendbaren Edelreiser sehr dünn sein müssen und damit die vorhandene Anwachsrate gering ist – wenig Energie im Edelreis. Die Methode sollte man jedoch auch weiter verfolgen!
Spaltung des Hypokotyls bis zur Radikula und dann Einlegen des Edelreises und verpacken mit Parafilm. Eventuell vorhandenen Austrieb abschneiden.
Abbildung 65: Ammenveredelung ins Hypokotyl: Edelreis u. Nuss li, vorbereitetes Edelreis und Nuss re
Abbildung 66: Ammenveredelung ins Hypocotyl: mit Parafilm li, plus Beschriftung re
Das obere Ende des Edelreisstiftes mit Baumwachs oder Parafilm abdecken!
Bei dieser Methode wird die Pfahlwurzel nicht beschädigt. Es können ewas dickere Edelreiser benutzt werden können als bei Veredelung ins Epilotyl, das Wurzelvolumen ist auch schon nach kurzer Zeit viel größer als bei der invertierten Radicula Methode (siehe Abschnitt 9.8.9). Die Anwuchsrate ist geringer als bei der invertierten Radicula Methode, jedoch entwickeln sich die Bäume durch die bessere Nährstoffversorgung später besser. Die Verbindung beider Pfanzen an der Veredelungsstelle lag bei dieser Methode auch bei nicht optimaler Anzuchtbedingungen (z.B. kein Gewächshaus) und auch Reiser alter Bäume im Mittel bei 50 %.
Einstiegsvariante:
Nach der Veredelung werden ein oder mehrere durchsichtige 3 l Säcke mit Zip vorbereitet. Zuerst den Sack beschriften und dann ca. ein Drittel des Sackes mit Torf befüllen. Die Nüsse mit den Reisern bis über die Schnittstellen in den lockeren Torf hinein drücken oder eingraben.
Hinweis: Bei der Verbindung über die Radikula ist die Nuss im Torf unten und die Radikula mit dem Edelreis zeigt nach oben.
Der Torf wird mit einer Blumenspritze etwas angefeuchtet und die Säcke zu drei Viertel verschlossen.
Die Säcke werden etwa eine Woche bei Raumtemperatur eher im Dunkeln, stehend gelagert.
Danach werden sie ca. drei Wochen bei Zimmertemperatur und bei Licht (Leuchtstofflampe weiß, 12 Stunden/Tag) zum Wachsen gebracht. Sinkt die Feuchtigkeit, so wird mit einer Blumenspritze nachbefeuchtet. Ab der dritten Woche werden die Säcke gänzlich geöffnet.
Verbesserte Einstiegsvariante:
Mittlerweile empfehle ich Anfängern leere 1,5 l PET-Einwegflaschen als „Gewächshaus“und auch als ersten Topf zu verwenden. Die Flasche in etwa 15 cm Höhe aufschneiden (Restverbindung belassen). Etwa 12 cm feuchten Torf locker einschichten und eine veredelte Nuss einsetzen. Das Halsstück mit wieder mit Klebeband fixieren und die Verschlußkappe abschrauben. Nach den Antrieb, das Halsstück entfernen und in den Boden der Flasche einige Entwässerungslöcher schneiden bzw. drücken.
Abbildung 67: Flaschengewächshaus
Eine Verbindung zwischen Amme und Edelreis durch Kallus braucht etwa 7–10 Tage. Nach etwa 3–4 Wochen gehören die Pflanzen aus den Plastiksäcken entnommen oder bei der verbesserten Einstiegsvariante die PET-Flaschen geöffnet. Bei den Methoden einer Veredelung direkt in die Nuss oder bei Veredelung an der Wurzelspitze, den Parafilm vorsichtig aufschneiden, damit sich Wurzeln ohne Hindernisse bilden können (siehe auch Abbildung 64).
Wurde die Pflanzen in Töpfe gesetzt, so kann man noch einige Tage eine Plastikhaube aufsetzen um die Luftfeuchtigkeit um die Blätter etwas zu erhöhen,
wobei eine Durchlüftung gewährleistet sein muss (Abbildung 72)!
Als weiteren Schritt werden die Pflanzen dann zur Abhärtung der Blätter im Halbschatten (Nord-Ost-Lage) ins Freie gestellt!
Professionellere Vorgangweise:
Eine geräumige Plastikbox mit minimal l x b x h = 35 x 25 x 25 cm oder beser l x b x h = 55 x 35 x 35 cm und durchsichtigem Deckel besorgen. Diese mit einem angefeuchteten Anzucht-Substrat mit Torf und Vermiculit (siehe auch 5.3 ) 10 cm besser 12 cm hoch füllen. Mit zwei Fingern Löcher drucken und die veredelten und beschrifteten Nüsse vorsichtig einsetzen. Die Deckelwände mit einer Blumenspritze leicht befeuchten und den Deckel verschließen.
Die gefüllte Box etwa eine Woche lang eher dunkel bei Raumtemperatur lagern, dann etwa zwei Wochen lang bei 12 Sunden Beleuchtung am Tag antreiben lassen und dann die Box ins Freie stellen. Zunächst den Deckel nur leicht öffnen und nach einer weiteren Woche ganz entfernen – hängt natürlich auch von der Außentemperatur ab. Bei Frostgefahr die Box in einem Raum stellen. Die Sonne nicht direkt auf die Box brennen lassen!
Abbildung 68: Ammenveredelung Anzuchtsack: noch ohne Torf li, mit Torf und stabilisiert re
Die Abbildung 68 zeigt li den Sack noch ohne Torf und re nach ca. 1 Woche in Anwuchsstellung mit Licht. Die Aufnahme wurden in der KW1/2015 gemacht und die re in der KW2/2015. Die Veredelung erfolgte zu früh. Im Jänner sollte nur dann veredelt werden, wenn ein richtiges Gewächshaus zur Verfügung steht.
Die Aufnahme der Abbildung 70 re wurde in der KW05/2015 gemacht und zeigt die ammenvermehrten Pflanzen im lockeren Torf.
Abbildung 72: Pflanztopf von 2015 li, mit Kappe zur kurzfristigen Erhöhung der Luftfeuchigkeit re
Abbildung 73: gut entwickelte Pflanzen li, Aufnahme KW12, Angesetzt KW9, keine Verbindung entstanden re
Abbildung 74: Versuch: Stieleiche als Amme – invertiert veredelt, gut verwachsen, über Winter abgestorben
Dre Monate nach der Ammenveredelung der dritten Versuchsreihe: Je größer die Ammennuss, desto besser ist das Wachstum der Pflanze. Starkwüchsige Hybridsorten wie Bouche de Bétizac entwickelten sich generell sehr gut. Das Edelreis kann man auch relativ spät schneiden – je nach Witterung – bis Anfang April. Die Knospen dürfen an den Spitzen grüne Ansätze zeigen, jedoch noch keine Blätter zeigen, sonst sinkt die Anwuchsrate drastisch! Der Anwuchserfolg bei Edelreisern, welche von Mitte Dezember bis zum Ende Februar geschnitten wurden, lag bei etwa 80 %. Bei Edelreisern, welche Ende März geschnitten und gleich eingesetzt wurden, lag die Anwuchsrate bei etwa 20–30 %. Die lange Lagerung der Ammennüsse, welche eine Spaltung von Stärke in Zucker, eine Verholzung bzw. Versprödung der Radikula bewirkt, wird einen nicht unwesentlichen Einfluss auf die Anwuchsrate haben. Interessanterweise wurde keine besonders gute Anwuchsrate zwischen Edelreis und Nuss vom gleichen Baum festgestellt. Nach dem Vereinzeln aus den Säcken in die Töpfe ist nochmals mit etwa 10 % Ausfall zu rechnen.
Ob sich die kleinen Bäume mit einem Alter von drei Monaten und 10–30 cm Größe später zum Aufschulen eignen, kann ich nicht abschätzen. Unveredelte Bäume (sat) erreichten im gleichen Zeitraum 20–50 cm an Größe, und unveredelte Hybriden (sat x cre) im gleichen Zeitraum eine Größe von 70–80 cm. Man muss auch berücksichtigen, dass mit einfachsten Mitteln gearbeitet wurde, dass also Licht, Temperatur, Luftfeuchte und auch die Nährstoffversorgung nicht an die Möglichkeiten einer Gärtnerei mit einem Gewächshaus herankommt.
Die Veredelung der Nüsse wurde im Zeitraum KW13 bis etwa KW 15 durchgeführt. Die Edelreiser wurden Ende KW04 geschnitten. Eine saubere Verbindung zwischen Edelreis und Nuss bei Bouche de Bétizac Reisern lag bei etwa 70 % vor. Bei anderen Sorten, wobei man auch meist eine geringere Reiserquualitätberücksichtigen muss, lag diese bei etwa 30 %. Wobei nochmals 20 % wegen mangelnder Wuchsleistung, vielleicht auch wegen zu kleiner Anzuchttöpfe abzuziehen sind.
Gut entwickelte Bäumchen besser gleich in 6 bis 11 Liter Töpfe mit seitlichen Drainangeschlitzen eintopfen (billige viereckige Töpfe zu diesem Zweck gibt es mit 19x19x23cm (6,5 l) 25x25x25,5 cm (11 l) von Cep und 18x18x23 cm (6 l), 23x23x26 cm (11 l) von Nuova Pasquini e Bini – NPB ). Damit bleibt die Substratfeuchte konstanter und die Wurzel können sich ungestört ausbilden. Im Untersetzer darf kein Wasser über längere Zeit stehen oder besser gleich keinen Untersetzer verwenden.
Wie man in den Bilden sehen kann, entwickelten sich nach der Veredelung im April 2017 bis zum Ende August 2019 einige 2 m hohe Bäume.
Abbildung 76: Ammenveredelung 2017: Anzuchtbox KW25/2017 li; ausgeleerter Inhalt KW25/2017 re
Abbildung 77: Ammenveredelung 2017: ausgeleerter Inhalt KW25/2017 li, im Topf KW30/2017 re
Abbildung 78: Ammenveredelung 2017: als Topfpflanzen KW27/2018 li, KW29/2018 re
Für den Kastanienrindenkrebs ist der Schlauchpilz (Cryphonectria parasitica) verantwortlich, Er befällt Kastanienbäume und Eichen, wobei er bei Eichen nur schwache Symptome bildet, und diese nicht zum Absterben bringt.
Der Pilz wurde einst mit asiatischen Kastanienbäumen nach Nordamerika eingeschleppt, wo die Krankheit erstmals 1904 im Zoo von New York/Bronx festgestellt wurde. Die asiatischen Kastanienarten Castanea crenata und Castanea mollissima bilden nur schwache Symptome aus. Der erste Import von Castanea crenata Bäumen in New York erfolgte laut [Pow98] durch S.B. Parson im Jahre 1875.
Innerhalb von 30 Jahren zerstörte er im östlichen Nordamerika einen Bestand von ungefähr 3 Milliarden Bäumen (American Chestnut – Castanea dentata) fast vollständig. Organisationen wie „The American Chestnut Foundation“ haben sich zum Ziel gesetzt, hoch-resistente Castanea dentata ähnliche Sorten zu züchten und damit eine großflächige Wiederaufforstung zu ermöglichen [Fou13].
In Europa wurde der Kastanienrindenkrebs erstmals 1938 im Hinterland von Genua entdeckt. Er breitete sich rasch im ganzen Mittelmeergebiet und dann über ganz Europa aus. Die Krankheit verläuft allerdings weniger dramatisch als in den USA.
Die von Wind verbreiteten Ascosporen oder vom Regen, Insekten, Vögel etc. verbreiteten Konidien gelangen durch kleine Wunden in die Bäume, wie etwa Rindenrisse, Astabbrüche, verlassene Gallen oder Veredelungsstellen. Der Pilz bildet ein gelbliches Mycel in der Rinde bis inklusive Kambium des Baumes. Er gehört zu den oxalsäurebildenden Schadpilzen. Das Toxin Oxalsäure greift das Kambium und den Bast (Phloem) – das Wachstums– und Transportgewebe des Baumes an und zerstört es. Der Schlauchpilz unterbricht damit den Transport von Nährstoffen wurzelwärts und das kambiale Wachstum. Erste Anzeichen für eine Erkrankung sind ein Einsinken oder ein Anschwellen des befallenen Gewebes sowie Risse in der Rinde. Glatte Rinde verfärbt sich rot. Später auftretende Symptome sind ein Welken der Pflanzenteile oberhalb der Befallsstelle und die Bildung von Trieben unterhalb der befallenen Stelle (Abbildung 82). Weiters sind welke Blätter bzw. im Winter trockene Blätter am Baum ein weithin sichtbares Symptom für einen Befall. C. parasitica bildet stecknadelgroße gebliche oder orange asexuelle und/oder sexuelle Fruchtkörper (Pyknidien und/oder Konidien). Die Abbildung 80 zeigt Fruchtkörper im Jänner. Die asexuellen Fruchtkörper Pyknidien entlassen bei feuchter Witterung die Sporen. In den sexuellen Fruchtkörpern (Perithezien) entwickeln sich in den Sporenschläuchen die sexuellen Sporen (Ascosporen). Die reifen Ascosporen werden herausgeschleudert und vor allem durch den Wind über Distanzen bis zu einigen hundert Metern ausgebreitet (??).
Der Pilz kann nicht in das Wurzelsystem eindringen, daher überlebt viele Wurzelstöcke und bilden wieder Stockausschläge.
In Europa gibt es insgesamt rund 64 VC-Gruppen (Vegetative Compatibility Groups) von Pilzen, wovon in Frankreich über 40 bekannt sind, in Italien, Bosnien und der Schweiz 20 bis 30, in Österreich 15, in Griechenland, wo alle Kastanienbestände befallen sind, allerdings nur 4. Die Anzahl der VC-Gruppen ist in den USA wesentlich höher.
Der bestehende, befallene Bestand an Bäumen wird in Europa mit hypervirulenten Pilzstämmen geimpft und so zu retten versucht. Projektleiterin für die Entwicklung eines Impfstoffes war Eva Wilhelm vom ARC Seibersdorf. In Österreich wurden 15 Pilzgruppen identifiziert, zwei davon – EU 17 und EU 13 – sind für 80 Prozent der Baumerkrankungen verantwortlich. Vor der Einfuhr von nicht zugelassenen Impfstoffen aus anderen Ländern wird gewarnt, kann es zu einer Erhöhung der VC-Gruppen kommen [Str06] [DR14]!
Abbildung 80: Kastanienrindenkrebs in Wien: Stamm mit Fruchtkörper li, Borke mit Fruchtkörper im Detail re
Abbildung 81: Kastanienrindenkrebs Stmk: herausgeschnitten teilweise vorhanden li, herausgeschnitten re
Anzumerken ist, dass Stämme von Cryphonectria parasitica in der Lebensmittelindustrie zur Käseerzeugung verwendet werden. Der Pilz sekretiert von Natur aus das Enzym Endothiapepsin, das die Milch zur Gerinnung bringt, indem das Enyzm spezifisch die Caseinmicellen hydrolysiert, und demnach nützlich für die Herstellung bestimmter Käse, insbesondere der Hartkäse wie des Emmentaler Käses ist [Bö03][SBI98]. Durch dieses Enzym wird das Labenzym Chymosin ersetzt. Früher wurde Chymosin aus dem vierten Magen von Kälbern gewonnen – nun wird es auch gentechnisch erzeugt. Gegenwärtig wird das Enzym Endothiapepsin in großtechnischem Maßstab durch Fermentationsverfahren erzeugt, die mit Stämmen von Cryphonectria parasitica durchgeführt werden.
Sofortmaßnahmen
Befallene Äste sollen unbedingt entfernt werden, um nicht als Infektionsherd zu dienen. Verletzungen jeglicher Art sind Eintrittspforten für den Pilz. Daher sind alle offenen Wunden mit Wundbalsam zu verstreichen.
Bei Veredelungen ist desinfiziertes Werkzeug und gutes Abdecken der Wunden wichtig, um den Befall zu vermeiden.
Jegliche Stressfaktoren wie Trockenheit, die den Baum schwächen, sind zu vermeiden [Kö15a].
Die Anlaufstelle in Österreich ist derzeit, 2015, die ARGE Zukunft der Edelkastanie [Sch14]. Man sollte ein Stück des befallenen Baumes labortechnisch prüfen lassen, um die Pilzstämme genau zu bestimmen und dann die Paste individuell herstellen lassen.
Resistente Pflanzen
Die japanische und chinesische Edelkastanie (Castanea crenata und mollissima) hat Resistenzen gegen den Schlauchpilz. Der Pilz kann sich bei diesen Bäumen nur oberflächlich ausbilden. Eine Bestrebung ist, durch Kreuzungen, die Eigenschaften wie Großwüchsigkeit der amerikanischen Edelkastanie (Castanea dentata) zu erhalten, d.h. letztendlich nur die Resistenzgene zu übernehmen. Das ist jedoch nicht einfach,
da insgesamt 3 Gene bei Castanea mollissima an der Resistenz beteiligt und diese auch nur teilweise dominant sind [K+13].
Ein anderer Weg zur Züchtung eines resistenten Baumes wurde durch Einbau eines Fremdgens (Transgene Pflanze) an der State University of New York eingeschlagen und steht vor einem Zulassungsantrag in den USA – 2015. Hier wurde ein natürliches Gen aus dem Weizen, welches oxalsäureabbauendes Enzym produziert, mit Hilfe des Bodenbakteriums Agrobacterium tumefaciens (indirekter Gentransfer) in das Genom der Castanea dentata eingebaut[ZNM+11]. Die Oxidase (OxO) zerlegt die vom Schlauchpilz ausgeschiedene Oxalsäure (C2H2O4) zu Kohlendioxid (CO2) und Wasserstoffperoxid (H2O2).
Für die Tintenkrankheit der Eelkastanie ist der Pilz Phytophthora cinnamomi und/oder Phytophthora cabivora verantwortlich. Es gibt über 60 verschiedene Arten von Phytophthora Pilzen. Die pilzlichen Erreger der Gattung Phytophthora zählen zu den gefährlichsten Schadorganismen an Pflanzen. Die sogenannten Oomyceten (Eipilze oder Scheinpilze) befallen vielerlei Nutzpflanzen und Bäume. Zu den wichtigsten Oomyceten im Ackerbau zählt Phytophthora infestans, der Erreger der Kraut- und Knollenfäule bei Kartoffeln. Zwischen 1816 und 1842 gab es 14 Kartoffel-Missernten, wobei der Grund für diese Serie vor allem auf den Ausbruch des Vulkans Tambora zurückzuführen ist, der weltweit das Klima beeinflusste, so dass das Jahr 1816 sogar als das Jahr ohne Sommer in die Geschichte einging. Der Dauerregen vernichtete den sandigen luftigen trockenen Boden, den die Kartoffel braucht, um optimal zu gedeihen. Im Jahr 1842 trat in Nordamerika erstmals die Kartoffelfäule auf. 1845 traten Ernteausfälle in den Niederlanden, Belgien und Frankreich und auch England auf. In Irland wurden nur zwei Kartoffelsorten angebaut, die beide anfällig waren. Somit fand der Oomycet in Irland besonders gute Bedingungen vor – er zerstörte ab 1845 die Kartoffelernten in Irland und löste dadurch eine Hungersnot beginnend von 1846 bis 1851 aus, die den Exodus von einer Million Iren zur Folge hatte.
Phytophtora megasperma löst bei Apfelbäumen die sogenannte Kragenfäule aus.
Phytophthora cinnamomi befällt die Wurzeln der Edelkastanie und anderer Pflanzen wie Eukalyptus, Eichen und auch Azaleen, Rhododendren, Zypressen, Wacholder, Heidelbeeren. Sein Temperaturoptimum liegt zwischen 24 und 28 ∘C.
Die Infektion geschieht an den feinen Wurzeln, über denen der Pilz ins Kambium größerer Wurzeln schließlich zum Stammgrund und Stamm weiterwandert. Der Baum hat schüttere Belaubung, die Blätter sind klein und oft vergilbt. Aus kranken Pflanzenteilen fließt bald ein schwarzer, dickflüssiger, tintenartiger Schleim. Die befallenen Bäume sterben nach 2–3 Vegetationsperioden ab.
Gefährdet sind Bäume mit einem geschwächten Wurzelwerk in staunassen Böden, aber auch nach starker Trockenheit, bei der viele Feinwurzeln absterben. Auch gegen diesen Pilz zeigen die asiatischen Edelkastanien Resistenz.
Maßnahmen
Fällen der betroffenen Bäume und der Nachbarbäume, Entfernen der Stöcke und des Wurzelsystems. Dabei muss darauf geachtet werden,
dass die Erde nicht verschleppt wird. Verbrennen von Wurzeln und befallenem Holz.
Infizierter Boden kann mit einem Fungizid gegen niedere Pilze (falscher Mehltau) behandelt werden (z. B. Ridomil) [Hei12]. Pflanzen, die mit Phosphonsäure (H3PO3) behandelt wurden, sollen sehr gut gegen Pilze aus der Gruppe der Oomyceten geschützt sein. Auch die Düngung mit Hühnermist soll Erfolge bringen.
Auf kontaminierten Objekten wie Pflanztöpfe lässt sich der Pilz durch 20 minütiges Tauchen in 60 ∘C heißes Wasser oder chemisch mit 2%iger Benzoesäure (Produkt MENNO Florades) bei einer Einwirkdauer von 4 Stunden entfernen.
Wird durch den Pilz Phloeospora castanicola (Septoria castanicola) hervorgerufen. Die Blattflecken, die dieser Pilz hervorruft, sind häufig zu finden. In den meist winzigen Flecken entwickeln sich Fruchtkörper und darin farblose Sporen, die weitere Blätter infizieren. Bei Massenauftreten kann die Krankheit vorzeitigen Blattfall auslösen. Da die Symptome meist keine Auswirkungen auf das Pflanzenwachstum haben, sind keine Maßnahmen notwendig.
Wird durch den Pilz Botrytis cinerea/Gnomoniopsis castanea hervorgerufen. Die Nüsse sind im Inneren verdorben, äußerlich ist der Befall meist nicht zu erkennen.
Früher Kastanienwickler, Buchenwickler, Esskastanienbohrer,
Blattläuse, Eichenblattlaus [Str06], Spinnmilben,
Thripse, Heuschrecken, Raupen, Maikäfer, Gallwespen.
Abbildung 83: Larve des Edelkastanienbohrers
Abbildung 84: Engerling 3 cm li, durch Wurzelfraß zugefügter Schaden an einer Topfpflanze re
Abbildung 85: Spinmilben auf Knospe li, Milben und deren Eier auf Blattunterseite re
Typische Blattfresser sind verschiedene Arten von Raupen (86) und Heupferde (87). In manchen Jahren kann es Wachstumseinschränkungen der Bäume durch massiven Blattfrass geben.
Abbildung 87: Grünes Heupferd auf Edelkastanienblatt li, Grünes Heupferd (Tettigonia viridissima) im Detail (4 cm) re
Viele Marienkäferarten sind Nützlinge, diese und deren Larven vertilgen Unmengen von Spinnmilben, Blättläuse etc.
Die Japanische Edelkastanien–Gallwespe (Dryocosmus kuriphilus) ist ein ursprünglich in Süd-China beheimateter Vertreter der Gallwespen. Sie gilt weltweit als größter tierischer Edelkastanien–Schädling. Die erwachsenen Tiere haben eine Körperlänge von 2,5–3 mm und sind schwarz (Abbildung 92). Es sind nur weibliche Tiere bekannt – die Vermehrung erfolgt parthenogenetisch (Nachkommen entstehen aus unbefruchteten Eizellen). Die weiblichen Wespen legen die Eier in Paketen von meist drei bis fünf, selten bis zu 30 Eier in die Blatt- und Blütenknospen der Kastanien. Insgesamt legt ein Weibchen über 100 Eier und stirbt dann nach wenigen Tagen. Die Eier sind 0,1 mm groß, oval geformt, weißlich, gestielt.
Die Larven schlüpfen und überwintern in den Knospen. Zu Beginn des Knospenaustriebs verursachen sie durch Ausschüttungen von Phytohormon–ähnlichen Wachstumsregulatoren die Gallen an den neuen Trieben (Abbildung 89). Die Larven fressen die nächsten 20 bis 30 Tage in der Galle, um sich dann zwischen Mai bis Juli zu verpuppen. Die Gallen sind 5 bis 20 mm groß und grün bis leuchtend rosafarben. Eine Galle enthält meist mehrere Gallwespenlarven. Sie verhindern eine normale Gewebeentwicklung. Die Blätter sind verkrüppelt, auch der Fruchtansatz wird großteils verhindert.
Ein verminderter Fruchtertrag mit Ausfällen bis zu 70 % sowie ein reduziertes Baumwachstum sind die Folgen. Es kann in Einzelfällen auch zum Absterben des Baumes kommen. Die von den Gallwespen gesetzten Verletzungen sind Eintrittspforten für den Edelkastanienrindenkrebs (Cryphonectria parasitica). Eine großräumige Verschleppung der Gallwespe ist durch den Verkauf befallener Bäume oder auch durch Äste wie Edelreiser möglich. Die natürliche Ausbreitung durch Flug beträgt etwa 8 km pro Jahr.
In Europa ist sie seit 2002 aus dem Piemont und seit 2005 aus Slowenien bekannt. In der Steiermark ist sie erstmals 2013 in Hausgärten, Plantagen und Waldstandorten aufgetreten. In Wien und Umgebung wurde sie von mir 2015 gesichtet. Die Gallwespe ist relativ kälteempfindlich.
Abbildung 89: Blatt mit Edelkastanien–Galle li, Edelreis mit Galle re
Abbildung 90: Schnitte durch Gallen
Abbildung 91: Gallenschnitt li, Larve 1,0mm re
Abbildung 92: Gallenschnitt mit entwickelten Gallwespen KW26/2015 li, flugfähige Wespe (3 mm) von unten re
Die Schlupfwespe, besonders die Schlupfwespe Torymus sinensis, ist ein natürlicher Feind der Gallwespe Dryocosmus kuriphilus [Qua15] [AA14]. Diese Wespen suchen die Gallen mit den Eiern der Gallwespe gezielt auf, stechen sie an und legen darin selbst ihre Eier ab, sodass die Gallwespen-Eier dem Schlupfwespen-Nachwuchs danach als Nahrung dienen und somit unschädlich gemacht werden. Wenn das Nahrungsangebot an Gallwespenlarven verschwindet, verschwinden auch wieder die Schlupfwespen.
Innerhalb der Gattung werden acht bis zwölf Arten unterschieden (Camus 1929) [Wik15d]:
| lat. Bezeichnung | beschrieben/kategorisiert | deut. Bezeichnung | eng. Bezeichnung | natürliche Verbreitung |
| Castanea sativa | Philip Miller 1754 (eng.) | Edelkastanie | European/Spanish C. | Europa und Vorderasien |
| Castanea dentata | (Marshall) Borkhausen 1800 | Amerikanische Kastanie | Chestnut | von Maine bis Alabama |
| Castanea crenata | Siebold u. Zuccarini 1846 | Japanische Kastanie | Japanese chestnut | Japan |
| Castanea mollisima | Blume 1851 | Chinesische Kastanie | Chinese chestnut | China |
| Castanea seguinii | Dode 1908 | Seguin Chestnut | China | |
| lat. Bezeichnung | beschrieben/kategorisiert | deut. Bezeichnung | eng. Bezeichnung | natürliche Verbreitung |
| Castanea ozarkensis | Ashe 1923 | Ozark chinkapin | Südstaaten von Amerika | |
| Castanea pumila | Miller 1768 | Allegheny chinkapin | Südstaaten von Amerika | |
| lat. Bezeichnung | beschrieben/kategorisiert | deut. Bezeichnung | eng. Bezeichnung | natürliche Verbreitung |
| Castanea henryi | Rehder u. E.H. Wilson 1916 | Henry chinkapin | in China | |
Die Castanea pumila wird manchmal in die Castanea ashei, floridana, alnifolia, paucispina (alle aus den Südstaaten) und pumilia aufgesplittet.
Die historisch maximale Größe der amerikanischen Castanea dentata ist unbekannt. Die Castanea dentata in Tervuren/Belgien wurde 1902 gepflanzt und war 2011 36,90 m hoch. Die europäische Castanea sativa erreicht ein Maximum von etwa 35 m (Studley Royal Deer Park/England – 2012 36 m, Wandstamm/Liebing/Österreich 2013 etwa 25 m). Gefolgt von der chinesischen Castanea mollissima mit etwa 15 m. Die japanische Castanea crenata erreicht etwa im Mittel 10 m.
Die Japanische Kastanie wird vom schwedischen Naturforscher Peter Karl von Thunberg in [Thu84] 1784 beschrieben. Sie wurde auch von Siebold und Zuccarini 1846 beschrieben und der Name Castanea crenata Sieb. et. Zucc. vergeben. Der Name Castanea mollissima Blume geht auf den Botaniker Carl Ludwig von Blume (1851) zurück. Die Castanea sativa wurde auch von dem deutschen Arzt und Botaniker Joseph Gaertner (1732-1791) beschrieben und als Castanea vesca Gaertn. bezeichnet.
Im deutschen Sprachraum wird noch gelegentlich die Bezeichnung Castanea vesca als Synonym für die Castanea sativa verwendet.
Anmerkung: Die Scheinkastanien (Castanopsis, engl. Chinkapin) sind eine Pflanzengattung in der Unterfamilie Quercoideae innerhalb der Familie der Buchengewächse (Fagaceae). Die etwa 120 asiatischen Arten sind im tropischen und subtropischen Gebieten verbreitet. Die Art Castanea henryi wurde 1899 von Sidney Alfred Skan als Castanopsis henryi erstbeschrieben und 1916 von Alfred Rehder und Ernest Henry Wilson in die Gattung Castanea gestellt. Früher wurden auch die Castanea ozarkensis und pumila der Gattung Castanopsis zugeordnet??
Von der Wuchsform kann man unterscheiden:
Bäume mit hoher kegelförmiger Krone wie Castanea dentata und sativa
Bäume mit breiter rundlicher Krone wie Castanea mollissima und ozarkensis
Sträucher mit halbkugeligen Wuchs wie Castanea pumilla und sequinii
Abbildung 94: Europäische Edelkastanie – Castanea sativa
Abbildung 95: Europäische Edelkastanie – Castanea sativa
Abbildung 96: Japanische Kastanie – Castanea crenata
Abbildung 97: Chinesische Kastanie – castanea mollissima
Abbildung 98: Castanea henryi
Abbildung 99: Castanea seguinii
Abbildung 100: Kastanienblättrige Eiche – Quercus Castaneifolio
Abbildung 101: Japanische Kastanien-Eiche – Quercus acutissima
Abbildung 102: Gesägte Eiche – Quercus serrata
Die Fossile Art der Castanea, die Castanea atavia Unger (1850) wird zeitlich dem Miozän (20,44-5,33 Mio Jahre) und dem Pliozän (5,33-2,6 MioJahre ) zugeordnet. Anmerkung: Die Gattung Castanea in diesem Zeitraum wird angezweifelt und derartige Blattformen in jüngster Zeit nun vermehrt der Gattung Quercus untergeordnet.
| Bezeichnung | Kurzbez. | Herk. | Kreuzung | Bestäuber | Anmerkungen |
| Belle épine | CA114 | FRA | sat | X | spätreif |
| Bouche Rouge | CA102 | FRA | sat | MS | spätreif, groß, guter Geschmack, haltbar |
| Bouche de Bétizac | CA125 | FRA | sat x cre | MS | frühreif, sehr groß, leicht schälbar, Gallwespen res., frostempf. |
| Bournette | CA112 | FRA | sat x cre | (MS) | mittelgroß bis groß, durchschnittlicher Geschmack |
| Castanea crenata | CA04 | JAP | cre | ||
| Castanea mollissima | CA75 | CHN | mol | X | Bestäuber für Hybride |
| Colossal | USA | sat x cre | MS | frühreif, gut schmeckend, frosthart | |
| Dorée de Lyon | CA111 | FRA | sat | MS | mittelreif, groß, sehr guter Geschmack |
| Ecker1 | AUT | sat | frühreif, mittelgroß, guter Geschmack | ||
| Ecker2 | AUT | sat x (cre x dent) | mittelreif, groß | ||
| Ferosacre | CA90 | FRA | sat x moll | frostempfindlich | |
| Maraval | CA74 | FRA | sat x cre | (MS) | spätreif, mäßiger Geschmack |
| Maridonne | CA124 | FRA | sat x cre | groß, wenig männliche Pollen | |
| Marigoule | CA15 | FRA | sat x cre | X | mittelfrüh, groß. durschn. Geschmack, gute Haltbarkeit, Holz |
| Marlhac | CA118 | FRA | sat x cre | MS | (F1) mittelreif, groß, gut schälbar, frosthart |
| Marron de Goujounac | CA500 | FRA | sat | X | mittelgroß bis groß, guter Geschmack, frostbeständig |
| Marron de Laguépie | CA109 | FRA | sat | groß, sehr süß, guter Geschmack, frostbeständig | |
| Marrone di segni | ITA | gut für Konditorwaren | |||
| Marsol | CA07 | FRA | sat x cre | X | mittelreif, groß, hoch empf. bei Gallwespe, Befr. für CA125 |
| Précoce Migoule | CA48 | FRA | sat x cre | frühreif, mittelgroß, gut schälbar, frostempfindlich | |
| Südtiroler Gelbe | ITA | sat | groß, sehr süß, leicht schälbar | ||
| Verdale | CA577 | FRA | sat | X | halb spät, mittelgroß, gut schälbar, frostbeständig |
sat – europäische Edelkastanie (Castanea sativa), cre – japanische Edelkastanie (Castanea crenata), moll – chinesische Edelkastanie (Castanea mollissima),
MS – männlich Steril, X – guter Befruchter.
Französische Hybridsorten (sat x cre) können einfacher von Stecklingen oder Abrissen vermehrt werden als es bei der europäischen Edelkastanie (Castanea sativa)
gegeben ist. Eine ausführliche Sortenliste ist unter [Ana] zu finden.
Bei [Cou12] werden Hybridsorten wie Marigoule, Marsol, Maraval, Marlhac vegetativ vermehrt, aufgezogen und nicht weiter veredelt. Sorten wie Belle épine, Bouche Rouge, Bournette, Bouche de Bétizac, Maridonne, Précoce Migoule werden auf Tintenkrankheit resistenten Unterlagen gezogen[Cou12].
Durch die Resistenz der C. crenata oder C. mollissima gegen Tintenkrankheit (Phytophtora) ist es naheliegend diese als Unterlage zu verwenden – es zeigten sich jedoch Inkompatibilitäten [PL04]. Genauer, laut einer nicht veröffentlichen Studie, welche in [Hua94] angesprochen wird, überlebten keine C. mollissima Veredelungen auf C. sativa Unterlagen. Und umgekehrt lag die Erfolgsrate bei C. sativa auf C. mollissima als Unterlage bei unter 20 % . Wobei eine typische Überwachsung der Unterlage durch das C. sativa Edelreis auftrat.
Genetische Inkompatibilität zwischen verschiedenen Sorten kann auch erst nach mehreren Jahren zeigen. Eine lokale Inkompatibilität zeigt sich an einer unzureichenden Ausbildung des vaskulären Verbindungsgewebes wie Xylem und Phloem über die Veredelungsstelle [PL04].
Rein europäische Sorten werden meist auf europäische Sämlinge veredelt. Alle Arten der Edelkastanie sind zueinander fertil [Hah04].
Es gibt Arten/Sorten, die untereinander genetisch gut kompatibel sind und manche schlechter. In [Cra92] wird eine genetische Analyse (Enzym-Elektrophorese) des Kambiumholzes verschiedener Fruchtsorten gezeigt und daraus eine Veredelungskompatibilität abgeleitet.
Genauer: Die Trockenmasse der Laub- bzw. Nadelhölzer enthalten 20 % – 35 % Lignin, der Rest besteht aus Cellulose und Chitin. Lignin ist ein
Biopolymer das eine Stützfunktion in der Pflanze hat – analog zu Glasfasern in Kunstharzen. Es entsteht biosynthetisch durch oxidative Polymerisation.
Den Polymerisationsprozess katalysieren extrazelluläre Peroxidasen aus Wasserstoffperoxid (H2O2) bzw. Laccasen (kupferhaltige Enzyme) aus Sauerstoff.
Die untersuchten Perioxidasen Enzyme des Kambium haben also bestimmende Rolle bei Veredelungsverbindungen. Enyzmatisch gleiche Gruppen erlauben ein unterbrechungsfreies vasculäres Leitgewebe (Xylem und Phloem). Die Studie [Hua94] mit einer chinesischen Sorte als Unterlage und verschiedenen Edelreisern
von C. dentata, crenata, mollissima widerspricht jedoch der aus der Perioxidase abgeleiteten Veredelungskompatibilität.
Empfehlungen zur Auswahl von Unterlagen und kompatiblen Veredelungen für französische Sorten nach [PL04] – Teilauszug:
| Unterlage | Res. Phytophtora | Res. Frühfrost | Kompatibilität | Wachstum | Kompatibilität |
| Ferosacre CA90 | *** | 0 | ** | 5 | |
| Belle épine | |||||
| Bouche de Bétizac | |||||
| Bournette | |||||
| Maridonne | |||||
| Migoule | |||||
| Précoce Migoule | |||||
| Marval CA74 | ** | ** | * | 2 | |
| Bournette | |||||
| Dorée de Lyon | |||||
| Maridonne | |||||
| Précoce Migoule | |||||
| Verdale CA577/CA756 | |||||
| Marigoule CA15 | *** | * | * | 4 | |
| Maridonne | |||||
| Précoce Migoule | |||||
| Marlhac CA118 | ** | * | ** | 3 | |
| Belle épine | |||||
| Bouche de Bétizac | |||||
| Dorée de Lyon | |||||
| Marigoule | |||||
| Migoule | |||||
| Verdale CA577 | |||||
| Marsol CA07 | * | ** | ** | 4 | |
| Belle épine | |||||
| Bournette | |||||
| Bouche de Bétizac | |||||
| Maridonne | |||||
| Marigoule | |||||
| Précoce Migoule | |||||
Eine noch größere Unsicherheit der Kompabilität ergibt sich meiner Meinung nach bei der Sämlingsvermehrung, nämlich auch dann, wenn der Sämling und Edelreis vom gleichen Baum stammen:
Der Baum wurde von einer anderen Sorte fremdbestäubt, der Sämling ist also ein gemischter Genotyp [Hua94],
welche Kompatibilitätsprobleme sind zu erwarten bzw. wie sind sie zu vermeiden?
Der Baum wurde z.B. selbstbestäubt, welche Kompatibilitätsprobleme sind zu erwarten?
Ist eine Baumgruppe genetisch gleich und reinerbig, so sind natürlich keine genetisch bedingten Kompatibilitätsprobleme zu erwarten!
Der pH-Wert ist ein Maß dafür, wie sauer oder basisch eine Lösung ist:
|
Die Abkürzung pH stammt aus dem lateinischen und steht für potentia hydrogeni – was Stärke des Wasserstoffs bedeutet.
Der angegebene oder gemessene Wert in wässrigen Lösungen ist der negative Zehnerlogarithmus der Wasserstoffionen–Aktivität (H+) in mol/l.
Auch in reinem Wasser entstehen durch Protonenaufnahme und Protonenabgabe:
2 H2O ⇌ H3O+ + OH− Oxoniumionen (H3O+ – sauer) und Hydroxidionen (OH− – alkalisch).
Der pH-Wert von Wasser liegt bei einer Temperatur von 25 ∘C bei 7. Bei 10 ∘C bei 7,27 und bei 40 ∘C bei 6,77!
Dieser Vorgang wird Autopyrolyse genannt, den es auch in Alkoholen, Schwefelsäure und Fluorwasserstoff und auch in Ammoniak gibt – näheres dazu siehe Abschnitt 17.
Zur groben Bestimmung des pH-Wertes kann man Wassertests aus der Aquaristik verwenden – Übersichtstests wie z.B. mit den „JBL pH-Test 3,0–10,0“. Achtung: beim Kauf auf den notwendigen pH-Wert-Bereich achten. Streifentests sind normalerweise noch etwas ungenauer.
| Will man den pH-Wert genauer bestimmen, dann sollte man mit einer pH-Einstabmeßkette arbeiten. Geräte wie das Voltcraft PHT-02 ATC gibt es relativ günstig zu kaufen. Diese pH-Meter haben jedoch den Nachteil, dass die Elektrodenmembranen bei Messungen von Bodenproben in wässriger Lösung relativ schnell verstopfen! |
Vorbereitung: Man nimmt zunächst einen Teil Substrat und vermischt es in einem sauberen Glas mit vier Teilen destilliertem Wasser, filtert nach einer halben Stunde die Mischung durch einen Filter (z.B. einem papierenen Kaffee- oder Teefilter) und wartet dann noch eine halbe Stunde um das Verschwinden der Trübung abzuwarten. Manche Bodenbestandteile wie Lehm setzen sich nur langsam ab. Bei einer Messung mit einer Farbindikatormethode und stärker gefärbtem Wasser, ergibt sich als Resultat eine Mischfarbe aus Umschlagsfarbe und Substrat, was natürlich den Vergleich mit einer Farbskala verfälscht.
Die nun angeführte Tabellen zeigen einige Meßergebnisse (pHH2O):
| Lehm | Floratorf | Lavakies 5-10 mm | Bims 15-20 mm | |
| Europomice | Europomice | |||
| Herstellerangabe | pHCaCl2: 2,5–3,5 | ph: 7–8 | ph: 7–8 | |
| JBL-Test | 6,5-7,0 | 4-4,5 | ||
| PHT-02 ATC | 6,5 | 4,2 | 7,15 | 7,2 |
| 1l FLoratorf+1TL Gartenk. | Substrat | Compo | Empfinger | |
| 5 g GK 14 Tage vor d. Messung | Topfpflanze | Guano | Aussaaterde | |
| Herstellerangabe | 70%ig | pH: 5,0–6,5 | ||
| JBL-Test | 6 | 5,5 | ||
| PHT-02 ATC | 6,5 | 6,8 | 6,7 | 6,75 |
Wobei zu beachten ist, welcher pH-Wert angegeben bzw. gemessen wurde.
Bei Messung in wässriger Lösung mit destilliertem Wasser, werden die an Bodenkolloiden (KAK) gebundenen H+-Kationen nicht mitgemessen (siehe auch Abschnitt 17.2.2), diese können jedoch aktiviert werden.
Bei einer Messung in Salzlösung von Calciumchlorid (CaCl2) oder Kaliumchlorid (KCl) kommt es zur Aktivierung der restlichen H+-Ionen.
Ursache: die leicht gebundenen Wasserstoffkationen (H+) werden von Calciumchlorid (CaCl2)- oder Kaliumchloridionen (KCl) verdrängt und sind dann in der Flüssigkeit frei beweglich und damit auch messbar. Dazu werden als Messlösungen 0,01 molare Calciumchlorid (CaCl2)- oder Kaliumchlorid (KCl)-Lösungen verwendet. Daher sollte bei einer pH-Wertangabe auf Substraten immer die Messmethode angeführt sein!
Will man den pH-Wert mit kohlensauren Kalk anheben, so ist auch feiner Gartenkalk mindestens eine Woche vor der Messung in das Substrat einzumischen und die Mischung feucht zu halten.
| Der gemessene pH-Wert mit destillierten Wasser als Lösungsmittel, der pHH2O, kann etwa 0,3−1,0 Einheiten über dem pHCaCl2 oder pHKCl liegen. |
Um notwendige Düngermengen zu bestimmen, Wasser zu enthärten, Insektizide zu verdünnen, oder auch Substratmengen zu bestimmen sind die Grundlagen über Einheitenumrechnungen, Mengenbestimmungen, Konzentrationsrechnungen und auch über chemische Verhältnisrechnungen notwendig.
Will man Einheiten sicher umrechnen, so sollte man von den Größenwerten ausgehen. Die Masse von 2,5 Kilogramm Dünger – Größenwert, schreibt man abgekürzt und formalisiert als m = 1,2 kg und allgemein als Ausdruck in der Form:
| Größenwert = Zahlenwert · Einheit G = | ⎧ ⎨ ⎩ | G | ⎫ ⎬ ⎭ | · | ⎡ ⎣ | G | ⎤ ⎦ |
Will man Einheiten wechseln, so muss der Größenwert zwischen alter Angabe und neuer Angabe gleich bleiben: malt = mneu; malt = 2,5 kg; mneu= 2500 g.
Für die Umrechnung von von 2,5 kg in g geht man folgendermaßen vor. 1 kg entspricht 1000 g man ersetzt Zahlenwert und Einheit:
| ⎧ ⎨ ⎩ | 2,5 | ⎫ ⎬ ⎭ | · | ⎡ ⎣ | kg | ⎤ ⎦ | = | ⎧ ⎨ ⎩ | 2,5 · 1000 | ⎫ ⎬ ⎭ | · | ⎡ ⎣ | g | ⎤ ⎦ |
Beispiel: Eine Düngeempfehlung mit 80 Pounds pro Acre Volldünger – in amerikanischen Maßeinheiten – soll auf g pro m2 umgerechnet werden. Dazu muss man zunächst wissen, dass 1 ac ≈ 4046,9 m2 und 1 lb ≈ 453,6 g entspricht. Dann überschreibt man im Ausgangsgrößenwert Zahlenwerte und Einheiten folgendermaßen:
| ⎧ ⎨ ⎩ | 80 | ⎫ ⎬ ⎭ | · | ⎡ ⎢ ⎢ ⎣ |
| ⎤ ⎥ ⎥ ⎦ | = | ⎧ ⎨ ⎩ | 80 | ⎫ ⎬ ⎭ | · | ⎧ ⎪ ⎨ ⎪ ⎩ |
| ⎫ ⎪ ⎬ ⎪ ⎭ | · | ⎡ ⎢ ⎢ ⎣ |
| ⎤ ⎥ ⎥ ⎦ | = | ⎧ ⎨ ⎩ | 8,97 | ⎫ ⎬ ⎭ | · | ⎡ ⎢ ⎢ ⎣ |
| ⎤ ⎥ ⎥ ⎦ |
80 lb/ac Dünger entsprechen also 8,97 g/m2.
Die genormten SI-Basiseinheiten (Système international d’unités) für Länge, Masse und Zeit sind Kilogramm, Meter und Sekunde (kg, m und s).
Für chemische Berechnungen wurde die SI-Einheit der Stoffmenge in Mol (mol) definiert. Ein Mol entspricht etwa 6,022 · 1023 Teilchen (Avogadro-Konstante). Praktischerweise hat ein Mol Wasserstoff etwa 1 g und 1 Mol Kohlenstoff etwa 12 g. Chemische Verhältnisrechnungen werden durch die Aufskalierung mit 6,022 · 1023 der atomaren Massen der Elemente vereinfacht! Für chemische Berechnungen wird die Atommasse (entspricht g pro Mol) des Periodensystems herangezogen. Im chemischen Periodensystem findet sich die Atommasse rechts über jedem Element. Mit einem einem Molmassenrechner wie [Van16] kann die Berechnung einfacher erfolgen.
Wie viel Massenprozente (w/w) Stickstoff enthält Ammoniumnitrat NH4NO3 . Aus der Theorie der Massenverhältnisse und einem Molmassenrechner lässt sich einfach mit
| NH4NO3 : 2· N = 80,043 : 2· 14,007 g/mol = 1 : 0,3499 |
berechnen. Ammoniumnitrat enthält also ungefähr 35 % Stickstoff (w/w).
Wie die Stoffmengenverhältnisse aussehen kann aus den Reaktionsgleichungen und Massenzahlen ausgerechnet werden.
Beispiel: Wie viel g reine Salpetersäure (HNO3) braucht man im Prinzip um 4 g Calciumcarbonat (CaCO3) durch eine chemische Reaktion in Calciumnitrat (Ca(NO3)2) überzuführen?
Die chemische Reaktionsgleichung dazu ist:
|
Das dazu benötigte Verhältnis von Calciumkarbonat zu Salpetersäure ist:
| CaCO3 : 2HNO3 = 100,087 : 2 · 63,013 mol/g = 1 : 1,259 |
Um 4 g Calciumcarbonat aufzulösen braucht man 4 · 1,259 = 5,036 g reine Salpetersäure.
( Bei einer 3 %gen (w/w) Salpetersäure sind in 100 g, 3 g reine Salpetersäure HNO3 enthalten. Man braucht 5,036 · 100/3 = 167,87 g 3 %ige Salpetersäure um 4 g Calciumcarbonat zu wandeln.)
Eine Lösung mit der Massenkonzentration c1 (weight to weight: w/w) und der Masse m1, soll in eine verdünntere Lösung mit der Konzentration c3 übergeführt werden. Anmerkung: Beim Arbeiten mit Volumskonzentrationen (v/v) sind natürlich auch die Massen durch Volumina zu ersetzen!
Ansatz über die Reinmengen – unverdünnte Substanzen:
|
|
Um die Gleichung zu erfüllen gibt es zunächst zwei Möglichkeiten: die Menge m1 wird fixiert und die benötigte Menge m2 ausgerechnet oder umgekehrt. Es ist aber auch möglich die Gesamtmenge mit m3= m1 + m2 vorzubestimmen:
Als notwendige Bedingung für positive Ergebnisse muss natürlich gelten:
| max(c1, c2) => c3 min(c1, c2) <= c3 |
Wird nur verdünndt wie mit destilliertem Wasser, so ist die Konzentration der c2 des Verdünners 0.
| m2 = m1 · (c1 − c3) / ( c3 − c2) |
Wieder der übliche Spezialfall: die Stoffmenge m1 wird mit einem Verdünner (wie z.B. dest. Wasser) verdünnt, daher ist dann c2 = 0.
Die zuzuführende Menge an Verdünner ist daher:
| m2= m1 · (c1/c3 −1) |
Beispiel: Aus einer 50 %igen Natronlauge (w/w) mit einer Masse von 100 g, soll eine Natronlauge mit einer Konzentration von 33,33 % hergestellt werden. c1 > c3 .. OK
| m1= 100 g, c1=50,0 %, c3= 33,33 % |
Einsetzen in die Formel
| m2 = m1 · (c1/c3 −1) = 100 · (50/33,33 − 1) = 50 g |
es muss 50 g destilliertes Wasser zugegeben werden.
| m3 = m1 + m2 = 100 + 50 g = 150 g |
Für den Fall, dass Menge 2 verdünnt wird: Vertauschen von 1 und 2!
|
Üblicher Spezialfall: Die Stoffmenge m1 in der Konzentration c1 wird mit einem Verdünner (wie z.B. dest. Wasser) m2 verdünnt, somit ist die Konzentration c2 des Verdünners also 0. Die obige Gleichung vereinfacht sich zu
| m1 = m3 · c3/c1 |
Beispiel: wie viel 50 %ige Natronlauge (w/w) und destilliertes Wasser (Verdünner) brauche ich für eine 10 % ige Natronlauge (w/w), wobei Gesamtmenge der verdünnten Natronlauge 100 g betragen soll? kleiner paranoia Check: c1 > c3 .. OK
Man braucht 20 g 50 %ige Natronlauge und 80 g destilliertes Wasser. |
Bei den Botenstoffen der Pflanzen, den Phytohormonen, handelt es sich um organischer Verbindungen, die eine ganze Reihe wichtiger pflanzlicher Entwicklungs- und Differenzierungsvorgänge wie Keimung, Wachstum, Samenreife, Blattabwurf, Blütenbildung, Zelldifferenzierung und Stammverzweigung steuern. Die fünf wichtigsten Pythohormongruppen sind Auxine, Cytokinine, Gibberelline, Ethylen und Abscisinsäure. Weitere Pythohormone sind Brassinosteroide, Salicylsäure, Jasmonsäure und Systemin.
Ihre Ausbreitung im Gewebe kann dabei entweder aktiv von Zelle zu Zelle erfolgen wie beim Auxin, über Leitbahnen (Phloem, Xylem) wie beim Auxin oder Cytokin, oder über den Gasraum zwischen den Zellen, wie beim Ethylen.
Mit den tierischen Hormonen teilen Phytohormone die Eigenschaft, schon in geringsten Konzentrationen wirksam zu sein und Signale auch zu weit entfernten Organen übermitteln zu können.
Im Gegensatz zu den Tieren haben Pflanzen jedoch keine echten deutlich abgegrenzten Hormondrüsen, sondern bilden sich je nach Entwicklungsstadien und Umweltbedingungen in verschiedenen Pflanzenteilen. Ein weiterer Unterschied zu den meisten tierischen Hormonen besteht darin, dass die Phytohormone nicht spezifisch auf ein bestimmtes Organ einwirken.
Auxine in der Sprosspitze, Meristeme, im embryonalen Gewebe über große Entfernungen im Phloem mit etwa 10 bis 20 cm/Std vom Spross in Richtung Wurzeln, über kürzere Strecken von Zelle zu Zelle Förderung der Adventivwurzel- und Seitenwurzelbildung, Kambiumaktivität, Streckenwachstum, Beteiligung an der Blütenentwicklung, Fruchtwachstum, Apikaldominanz, verhindert Blatt- und Fruchtabwurf; Transport immer von der Sprossachse immer abwärts und in den Wurzeln immer zur Spitze. Cytokinine Wurzelspitzen, keimende Samen, wachsende Gewebe mit Proteinsynthese von den Wurzel zum Spross im Xylem Zellteilung und Differenzierung, inkl. Entwicklung von Kallus, Chloroplastenentwicklung, Verzögerung von Alterungsprozessen, fördert Seitenknospen Gibberelline im jungen Gewebe: unreife Samen und Früchte, Embryonen und Pollenkörner wahrsch. im Xylem und Phloem Sprossverlängerung durch Streckungswachstum (Zellstreckung), Blütenbildung, Geschlechtsausbildung bei monözischen Pflanzen Abscisinsäure Blätter und Wurzeln übers Xylem von Wurzel und Phloem von dem Blättern Schließung der Respirationsspalten bei Wassermangel, Auslösung der Dormanz: verhindert vorzeitige Samenkeimung, Knospung Ethylen in den meisten Geweben als Reaktion auf Stress, stark in reifenden Früchten als gasförmiger Stoff in den Interzellularen, aber auch zwischen verschiedenen Pflanzen Einleitung von Ruhestadien, Förderung der Fruchtreife und Alterungsvorgängen, fordert die Blütenbildung und des Laub- und des Fruchtfalls
Die Apikaldominanz, die Dominanz der Endknospe am primären Spross, verhindert über eine Auxinausschüttung das Wachsen von Seitentrieben.
Sie ist besonders bei Pflanzen ausgeprägt, die im Schatten anderer Pflanzen wachsen und eine Schattenflucht aufweisen. Pflanzen mit starker Apikaldominanz sind beispielsweise Sonnenblumen, während die
Apikaldominanz bei Paradeisern nur gering ausgeprägt ist.
Buchengewächse wie die Edelkastanie haben angeblich nur eine geringe Apikaldominanz, da die Edelkastanie über den Winter die apikale/terminale Knospe verliert. ??
Auxine, welche in der Terminalen Knospe gebildet werden, werden in basaler Richtung transportiert, bewirken auch über größere Entfernungen hinweg die Unterdrückung der Seitentriebentwicklung im unteren Teil der Pflanze. Diese vollständig oder auch teilweise ablaufende Unterdrückung von Seitenknospen wird vor allem durch IAA vermittelt. Schneidet man das terminale Ende des Hauptsprosses weg, so werden sich Seitentriebe ausbilden.
Die Dormanz von Samen, welche durch Abscisinsäure gebildet wird, verhindert die Keimung von Samen in der Frucht und auch die Keimung in der Winterperiode. Mit Gibberellinen kann man jedoch die Dormanz bei vielen Samen brechen und sie dadurch zur frühzeitigen Keimung ohne Kälteperiode oder Licht bringen. Gibberelline können bei manchen Pflanzen Schossen und Blütenbildung auslösen.
Für die Differenzierung des Zellgewebes bzw. Organbildung ist das Mengenverhältnis von Auxin (IAA) zu Cytokinin (Kinetin) und die absolute Menge entscheidend. Relativ hohe IAA und Kinetin Konzentrationen führen z.B. bei Stängelmark von Tabakpflanzen in Gewebekulturen nach einigen Wochen zur Bildung von Kallus hier beispielhaft mit IAA : Kin = 3 : 0,2 mg/l. Bei Erhöhung des Verhältnisses IAA : Kin = 3 : 0,02 mg/l entwickeln sich ausgehend vom Kallusgewebe nur Wurzeln. Bei einer Umkehrung des Verhältnisses IAA : Kin = 0.003 : 1 mg/l entwickeln sich aus dem Kallus nur Sprosse. Und bei einer Gabe nur von Kinetin IAA : Kin = – : 0, mg/l bildet sich kein Kallusgewebe und damit auch keine Organe aus dem Stängelstück.
| Bei einem hohen Auxin zu Cytokinin-Verhältnis bildet sich Wurzelgewebe, bei einem niedrigen Auxin zu Cytokinin-Verhältnis bildet sich ein Spross. Diese Wirkung wird in pflanzlichen Gewebekulturen ausgenutzt. In der Pflanze kommt es bei hoher Auxinkonzentration zu vermehrter Bildung von Adventiv- und Seitenwurzeln. Außerdem wird im Kambium bei einem Auxin-Cytokinin-Verhältnis von ungefähr 1:1 Xylemgewebe gebildet. Das spielt vor allem in der Pflanzenentwicklung und nach Verwundung eine Rolle. Bei hohen Konzentrationen von Auxinen werden Wurzel- und Sprosswachstum gehemmt, da die Produktion von Ethylen gefördert wird. |
|
Die Bewurzelungshilfen bewirken keine Wunder und wurden bei der Ammenvermehrung nicht verwendet. Mit den angeführten Phytohormonen sollte man zur eigenen Sicherheit
vorsichtig umgehen. Generell wird empfohlen Gummihandschuhe, Schutzkleidung und dichten Augenschutz zu verwenden, Staubbildung und das Einatmen der Substanzen ist zu vermeiden! Restmengen und nicht wieder verwertbare Lösungen einer Problemstoffsammelstelle (in AT gem. ÖNORM S2100) übergeben. 4-(Indol-3-yl)buttersäure (IBA) wird nach dem Global Harmonisierten System (GHS) als giftig (GHS06) eingestuft. Zutreffende Codes sind: H301, H315, H319, H335 – giftig bei Verschlucken, verursacht Hautreizungen, verursacht schwere Augenreizungen, kann Atemwege reizen. Genauere Angaben zur Handhabung von IBA findet man in den Sicherheitsdatenblättern von [Rot13] oder [SA14]! |
In höheren Pflanzen kommt das Auxin Indol-3-essigsäure (IAA) neben anderen Phytohormonen in geringer Menge von 1 bis 100µ g je kg Pflanzenmaterial vor. Die 4-(Indol-3-yl)buttersäure (IBA) wurde aus Mais und Weiden isoliert.
Bei einer zu hohen Konzentration von IAA wird die Synthese von gasförmigem Ethylen gefördert, einem Stress-„Hormon“, welches sich beispielsweise negativ auf das Längenwachstum der Wurzeln auswirkt. Kommerziell wird IAA meist nicht verwendet, da sie in wässriger Lösung durch eine Spezies von Acetobacter [HK14] und durch Licht relativ schnell abgebaut wird und daher nur gekühlt und dunkel etwas länger haltbar ist. Die Acetbacter haben keinen Effekt bei IBA–Lösungen. Bei kristalliner IAA ist angeblich bei einer Lagertemperatur von −20 ∘C keine nennenswerte Veränderung über eine längere Zeit hinweg zu erwarten.
Bewurzelungshilfen mit Algenpräparaten sind nicht das gleiche – da kann man gleich ein Präparat aus Weidenwasser ansetzen!
Die meisten stark verdünnten wässrigen Lösungen von Phytohormonen werden von Mikroorganismen rasch abgebaut und sollten daher auch gekühlt gelagert nur 1–2 Wochen lang verwendet werden. Eine Zusammenstellung der Auxinapplikation zur Bewurzelung von Stecklingen findet man in [BSTR07].
Es werden am Markt diverse fertige Bewurzelungshilfen in Pulver-, Tabletten- und Gelform angeboten. Die bekannteste Marke in Europa (NL), welche auch eine ausführliche Homepage hat, ist Rhizopon. Die Firma vertreibt verschiedene Bewurzelungshilfen wie Rhizopon AA Tabletes (Tabletten mit 0,5 % IBA zum Auflösen in Wasser), Rhizopon AA Poeder (0,3 % IBA in Puder), Rhizopon Dip’n Grow (1,0 % IBA 0,5% NAA). Rhizopon IBA Produkte sollen 2016 in den Niederlanden neu zugelassen werden. Andere Präparate werden unter Handelsnamen wie Clonex Rooting Gel (0,3 % IBA, 0,5 % Benzisothiazolinon als Mikrobizid und Fungizid, 2-Hydroyxethylcellulose als Gelbildner, Propylene Glycol und Wasser), Sangral Powder (NAA und Thiram als Fungizid), Dueci Fito Radicante (0,5 % NAA im Talkum) von Guaber – 2013 in IT registriert, Seradix (B1 – 0,1 % B2 – 0,3 % B3 – 0,8 % IBA im Talkum) von Kwizda Agro vertrieben – ist in AT bis 31.5.2015 zugelassen (Abverkaufsfrist: 31.5.2016). Von Phytronics wird das Produkt Hortus IBA Water Soluble Salts (20 % IBA und einen pH Puffer), welches in den USA EPA (Reg No. 63310-22) zugelassen ist, vertrieben. Das Salz kann bis zu einer Konzentration von 6 % IBA in Wasser gelöst werden. Für Auxine und Anwendungen siehe auch [HK14][Bel08][SA15].
Die Rezepturen sind aus dem Internet zusammengestellt und setzen einen professionellen Umgang mit den teils giftigen Ausgangssubstanzen voraus. Da 4-(Indol-3-yl)buttersäure (IBA) mit CAS 133-32-4 laut CLP-V (Classification, Labelling and Packaging-Verordnung) als giftig mit GHS 06 ausgewiesen wird, darf es in reiner Form nur an Forschungsinstitute, Universitäten und Berechtigte mit Gewerbeschein abgegeben werden, daher sind Bewurzelungsrezepturen im folgendem Text mit IBA ausgegraut. Abgesehen davon sollte jeder selbst abwägen, ob er eine Bewurzelungshilfe mit reinem IBA selbst herstellen möchte! Alternativ können fertige Bewurzelungshilfen oder auch oben angeführte wasserlösliche Tabletten oder Salze als Auxinquelle für Rezepturen eingesetzt werden - auch mit diesem ist sorgfältig umzugehen!
Auch beim Mischen von Kleinstmengen sollten Einweggummihandschuhe, Labormantel und eine abgedichtete Schutzbrille verwendet werden. Neben labortechnischen Gerätschaften wie Messbecher, Mörser, Löffel, Mixer, Folien, Abfüllbehältnisse, Thermometer und Spachteln wird man um eine hochpräzise Taschenwaage für die Wägung von Kleinmengen nicht hinwegkommen. Vorteilhaft sind Taschenwaagen mit einem Wägebereich bis 200 g/300 g und einer Ablesbarkeit oder besser einer Genauigkeit von 0,01 g. Das Kalibriergewicht sollte dabei sein!
Anmerkung: Die meisten Auxine sind in Wasser nur schwer löslich und müssen in Alkohol oder einer Lauge vorab aufgelöst werden. Die Kaliumsalze (K-IAA, K-IBA) der Säuren sind direkt im Wasser lösbar aber auch teurer.
| Abk. | CAS-Nr | Name | LagerT. Pulver | gut Löslich in | Anmerkung |
| IAA | 87-51-4 | Indol-3-essigsäure | −20 ∘C | Natronlauge, Ethylalk. (100–1000 g/l), Isoprop. | instabil, lichtempf. |
| NAA | 86-87-3 | 1-Naphthylessigsäure | Raumtemp. | Natronlauge, Aceton, Ethylalk., Isopropanol | stabil, etwas lichtempf. |
| IBA | 133-32-4 | 4-(Indol-3-yl)buttersäure | +2 − +8 ∘C | Natronlauge, Ethylalk. (30–100 g/l), Isoprop. | stabil, laut CLP giftig |
| K-IAA | 2338-19-4 | Kaliumsalz-Indol-3-essigs. | +2 − +8 ∘C | Wasser | |
| K-IBA | 60096-23-3 | K-4-(Indol-3-yl)butters. | +2 − +8 ∘C | Wasser | |
Gefahrenhinweise und Löslichkeiten für mehrere Wachstumsregulatoren sind auf Sigma Aldrich [SA15] zusammengestellt.
Das basale Ende des Stecklings mit etwa 1/5 seiner Länge für 2–10 Sekunden in das Bewurzelungsmedium eintauchen. Den Steckling gleich danach einsetzen.
|
1,0 g IBA in 20 g Ethylalkohol, Präparat mit 4,8 % IBA (w/w) als Stammlösung 1,0 g IBA Indol-3-essigsäure in 20 g (etwa 25 ml) Ethylalkohol (Spiritus geht natürlich auch) auflösen.
Die angesetzte Stammlösung ist in einem dunklen Glasfläschchen gekühlt etwa ein Jahr haltbar. |
Anmerkung: Ethylalkhohol hat eine Dichte von etwa 0,8 g/ml. Die Konzentration in % der Wirksubstanz (IBA) in der obigen Stammlösung wurden als Massenprozente (w/w) gerechnet, also Masse IBA durch Gesamtmasse mal 100.
|
50 ml IBA-Alkohol-Wasser-Lösung mit 0,5 % IBA (w/w) und etwa 25 % Ethylalkohol (w/w) 4,7 g (5,88 ml) der obigen IBA-Ethylalkohol-Stammlösung enthalten 0,235 g IBA. Diese mit 9 ml Ethylalkohol und 35 ml destilliertem Wasser verdünnen. Probe: 0,235/(14,88 · 0,8+35) · 100 = 0,501 % Anmerkung für die Praxis: Ein 1/4 der Stammlösung in eine 50 ml Flasche füllen, diese mit Ethyalkohol auf etwa 1/3 auffüllen und dann den Rest mit destilliertem Wasser ergänzen. |
100ml Präparat mit 1,5 % IAA (w/w) als wässrige Lösung
1,5 g IAA Indol-3-essigsäure (C10H9NO2) in 2 ml Isopropanol oder Ethylalkohol (Spiritus geht auch) oder besser
in 1 molarer Natronlauge auflösen und diese Lösung mit 100 (99) ml destilliertem Wasser verdünnen. Lösung ist auch gekühlt nur kurz lagerbar
|
0,5 % IBA und 0,25 % NAA in 100 ml dest. Wasser, Präparat als wässrige Lösung 0,5 g IBA und 0,25 g NAA in 2 ml 1 molarer Natronlauge auflösen und diese Lösung mit 98 ml destilliertem Wasser verdünnen.
Die so hergestellte Bewurzelungsflüssigkeit ist gekühlt und abgedunkelt max. 2 Wochen lagerbar. |
Etwa 1 cm über dem basalen Ende des Stecklings ein Loch quer durch den Steckling mit einer Stechahle oder Bohrer in Zahnstocherstärke machen und den präparierten Zahnstocher durch dieses Loch stecken. Der Zahnstocher muss fest mit dem Steckling verbunden sein.
|
10ml wässriges Präparat mit 2 % IBA und 1 % NAA 0,20 g IBA und 0,10 g NAA in etwa 5 Tropfen 1 molarer Natronlauge auflösen und diese Lösung mit 9 ml destilliertem Wasser verdünnen.
Die so hergestellte Bewurzelungsflüssigkeit in ein mindestens zahnstocherhohes Fläschchen abfüllen. |
Runde Zahnstocher in das Fläschchen mit der Bewurzelungsflüssigkeit geben, das verschlossene Fläschchen schütteln und die Zahnstocher 24 Stunden darin quellen lassen. Die Zahnstocher mit einer Pinzette herausnehmen und etwas antrocknen lassen. Will man die präparierten Zahnstocher lagern, dann diese in ein trockenes beschriftetes dunkles Glasfläschchen einschlichten und dieses einfrieren.
Vorteile: Die Auxine werden langsam in den Steckling mit wenig Schwund eingebracht. Die benötigte Menge an Auxinen ist bei dieser Methode sehr gering!
Für eine 1 molare Natronlauge (Molekulargewicht 40) löst man 40g Ätznatron in einem Liter destilliertem Wasser auf – oder entsprechend 1 g Ätznatron in 25 ml dest. Wasser.
Will man die wässrigen Lösungen als Gießwasser verwenden oder Stecklinge ins Wasser stellen, so sind Konzentrationen in der Größenordnung
1: 107 für IAA : Wasser angebracht (0,1 ppm IAA):
0,01 % IAA in 100 g, Präparat als wässrige Stammlösung
0,010 g IAA Indol-3-essigsäure (C10H9NO2) in 1 ml Isopropanol oder Ethylalkohol oder 1 molarer Natronlauge auflösen
und diese Lösung mit etwa 98 ml destilliertem Wasser verdünnen (Stammlösung).
Die hergestellte Stammlösung ist auch im Kühlschrank nicht lange lagerfähig
0,3 ppm IAA als Gebrauchslösung
3 ml der obigen Stammlösung in 1 l Wasser einmischen.
Weidenwasser
Zur Herstellung junge Weidentriebe mit grüner oder gelber Rinde abschneiden und die Blätter entfernen. Die Rinde durch klopfen lockern und abziehen, dann die Rinde und das Holz in 1–2 cm Stücke zerschneiden und in ein Glas mit Deckel füllen. Zum Schluss das Glas mit kaltem Wasser auffüllen und locker verschließen. Den Ansatz 2–3 Tage an einem kühlen Platz oder Kühlschrank ziehen lassen und abseihen, und dann die Flüssigkeit im Kühlschrank aufbewahren. Diese Lösung kann man ins Gießwasser gegeben oder Stecklinge darin eintauchen. Anmerkungen: Die in der Weide enthaltene IAA/IBA??, Gerbstoffe, Phenolglykoside, Salicin und Salicinderivate werden im Wasser gelöst. Problematisch dürfte auch die rasche Zersetzung von IAA durch Bakterien bei einem Kaltauszug sein.
Den Steckling anfeuchten und das basale Ende eines Steckling etwa 1/5 seiner Länge in das Bewurzelungsmedium eintauchen und danach den Steckling einsetzen.
100 g Pulverpräparat mit 0,5 % NAA
0,5 g NAA in 50 ml Aceton auflösen. Dann 100 g Talkumpuder sorgfältig einrühren
und das Gemisch anschließend trocknen lassen.
Das Pulver wird abgefüllt und beschriftet.
Das so hergestellte Bewurzelungspulver ist bei Raumtemperatur 1–2 Jahre lagerfähig
20g Pastenpräparat mit 0,5 % IAA für Stecklinge
0,5 g x 2 / 5 = 0,2 g Kaliumsalz IAA in 10 ml destilliertem Wasser auflösen und
10 g wasserlösliches Wollfett in 10 ml Lösung sorgfältig einrühren.
Die hergestellte Paste ist nicht lagerfähig
|
100 g, Präparat als Paste mit 0,4 % NAA und 0,4 % IBA [dIFdL14] für Abrisse 0,4 g NAA + 0,4 g IBA in 10 ml Ethylalkohol einrühren |
Die Edelkastanie ist ein Kernholzbaum welcher im trockenen Zustand leicht zu bearbeiten ist. Im Freien sind Bretter und Pfähle auch ohne chemische Behandlung weitgehend witterungs- und fäulnisbeständig, da es einen hohen Gehalt an Tanninen enthält. Das Holz trocknet nur langsam und neigt zum Reißen und Werfen, daher muss die Trocknung sehr langsam durchgeführt werden.
Das schmale, oft nur 2 bis 5 Jahresringe breite äußere Splintholz ist gelblich-weiß gefärbt, das Kernholz von gelbbrauner bis dunkelbrauner Farbe. Das Kernholz dient als Totholz als zusätzliche
Stütze des Stammes. Der Baum hat ein typisch ringporiges Holz. Die Frühholzporen sind sehr groß und auffällig oval; Durchmesser bis 0,5 mm (Xylem) in radialer Richtung (Abbildung 106 re). Im
Längsschnitt des Stammes/Äste bilden Längsporen (Xylemgewebe) deutliche Nadelrisse (Abbildung 106 li). Im Gegensatz zur Eiche sind die radialen Poren mit freiem Auge fast nicht sichtbar.
Rohdichte bei ca. 0 % Holzfeuchte 530–590 kg/m3
Härteklasse: mittelhart
Schwindmaß in Längs–/radialer Richtung/Volumen: 0,6/4,3/11,5%
Abbildung 106: Längsschnitt li, Querschnitt re
Abbildung 107: Mikroskopischer Quersch. mit Borke: Färbung Etzold FCA li, ungefärbt re
Die Etzold FCA-Färbung (Fuchsin, Chrysoidin, Astrablau) [Aei19] färbt verholzte (lignifizierte) Zellwände in verschiedenen Rottönen. Cutinisierte Zellwände werden gelb, unverholzte Zellwände werden blau gefärbt. Die Borke bleibt nahezu ungefärbt. Eingebettet wurde mit Eukitt von Kindler [Kin19].
| Pflanzen erzeugen energiereiche Stoffe mit Hilfe von Licht und der Photosynthese aus energiearmen Stoffen wie Kohlenstoffdioxid (CO2) aus der Luft und Wasser (H2O). Über mehrere Reaktionsebenen wird Sauerstoff (O2) und energiereiche Glucose (C6H12O6) erzeugt. |
Pflanzen benötigen neben Wasser und Kohlenstoffdioxid weitere Nährstoffe wie Stickstoff (N) um die stickstoffhaltigen Aminosäuren und Nucleinsäuren zu bilden. Unsere Luft besteht aus rund 78 % Stickstoff. Die Aufnahme des Stickstoffs erfolgt leider in der Regel durch Nitrat-Aufnahme aus dem Boden und nicht direkt aus der Atmosphäre. Es gibt aber auch Pflanzen, die den Luftstickstoff indirekt durch eine Symbiose von Bakterien mit der Pflanzenwurzel aufnehmen können – dazu zählen Hülsenfrüchtler (Leguminosen) wie Erbsen, Bohnen (Ackerbohnen), Linsen, Klee (Rotklee, Weißklee, Perserklee) und auch Bäume wie Rot-Erle (Alnus rubra), Grau-Erle (Alnus incana) und Ölweiden (Elaeagnus).
So gehen Hülsenfrüchtler dazu eine Symbiose mit speziellen Bakterien (Rhizobien) ein, die zu Ausbildung von Wurzelknöllchen führen und für sie molekularen Stickstoff umwandeln [Kri10].
Rhizobien besitzen die Fähigkeit, elementaren, molekularen Stickstoff (N2) zu binden, indem sie ihn zu Ammoniak (NH3) bzw. Ammonium (NH4+ ) reduzieren und damit biologisch verfügbar machen. Dies ist ihnen jedoch nur in der Symbiose mit Pflanzen möglich.
Bei Bäumen kann die Stickstoffixierung mit dem Bakterium Frankia alni 240 bis 350kg/ha und Jahr betragen. Klee kann 50 bis 350 kg/ha Luftstickstoff fixieren.
Freilebenden Bakterien wie Azotobacter, Azomonas und Cyanobakterien können etwa 1–3kg/ha Luftstickstoff fixieren.
Neben dem Austausch der Grundelemente Kohlenstoff (C), Sauerstoff (O) und Wasserstoff (H), die in Kohlenstoffdioxid (CO2) und Wasser enthalten sind, gelten nur 13 dieser Elemente als lebensnotwendige Nährelemente, welche im Regelfall über die Wurzeln aufgenommen werden.
Die Einteilung der benötigten Nährstoffelemente geschieht nach der benötigten Menge in:
Makronährelemente sind Stickstoff (N), Phosphor(P), Kalium(K), Schwefel (S), Calcium(Ca) und Magnesium(Mg)
welche über Wurzeln aufgenommen werden.
Mikronährelemente oder Spurenelemente werden nur in geringen Mengen benötigt, diese sind Mn, B, Zn, Cu, Mo, Cl und Fe. Eisen (Fe) ist unter anderem für die Blattgrünbildung verantwortlich. Das typische Eisenmangelsyndrom ist die sogenannte Christbaumchlorose, bei der sich die Blattflächen gelblich sind, während die Blattadern grün sind (siehe auch Abschnitt 17.5.4).
Die Düngefibel [Hau12] von Hauert, einem Düngemittelhersteller für Gärtnereien, bietet in Kompakter Form mit viel Zahlenmaterial im Hintergrund, einen praxisbezogenen Leitfaden für die richtige Düngung.
Voraussetzung für die Wasseraufnahme der Wurzel ist, dass die Salzkonzentration in den Wurzelzellen höher ist als im Bodenwasser.
Düngersalze müssen somit bei der Aufnahme in einen Bereich hoher Salzkonzentration wandern. Da dies nicht möglich ist, müssen die Ionen unter Energieverbrauch gegen ein Konzentrationsgefälle in die Wurzelhaarzellen geholt werden. Da die Zellmembran für die Ionen undurchdringlich ist, werden sie von speziellen Trägern hindurch transportiert und gelangen so ins Zellinnere.
Pflanzen können Nährsalzionen selektiv aufnehmen. Das liegt daran, dass für bestimmte Ionen bestimmte Träger vorhanden sind, die sich an der elektrischen Ladung und dem Ionendurchmesser orientieren. Das Kalium- und das Ammoniumion besitzen die gleiche Ladung und ungefähr den gleichen Durchmesser. Aus diesem Grund konkurrieren sie um den gleichen Träger. Herrscht ein Überangebot an einem der beiden Ionen, wird das anderem in geringerer Menge aufgenommen. Dieses Phänomen kann zu einem Nährstoffmangel führen und nennt sich Antagonismus. Diese Erscheinung tritt beispielsweise auch in gut mit Kalk versorgten Böden oder Substraten auf. Dort sammeln sich die Calcium-Ionen aufgrund ihrer großen Konzentration in der Bodenlösung an der Wurzel an und hemmen die Aufnahme anderer Ionen, wie zum Beispiel Magnesium- und Kalium-Ionen!
Genauer betrachtet [Wik15e] und trotzdem stark vereinfacht, geschieht die Aufnahme der Nährstoffe (Ionen) über selektive Diffusion über ein elektrisches Potentialgefälle gegen das Konzentrationsgefälle und einen aktiven Transport durch die Wurzelzellmembranen.
Wurzeln können schwach gebundene Kationen wie Ca2+, Mg2+, K+, Na+, NH4+ in Lösung bringen oder auch schon gelöste über die Wurzeln assimilieren. Genauso können Wurzeln Anionen wie PO43−, SO42−, Cl−, NO3− aufnehmen.
Wobei immer auch Ionen von den Wurzel abgegeben oder dissoziiert werden um die Ladungsneutraliät zu erhalten.
Die Kohlensäure-Austausch-Theorie [Elh14] ist das bekannteste Modell: die Wurzeln geben Kohlenstoffdioxid (CO2) ab und in Wurzelnähe im Wasser des Substrates dissoziiert es zu:
|
Wird ein Kaliumkation (K+) von der Wurzel aufgenommen, so wird es gegen ein Proton (H+) eingetauscht. Wird ein Chloridanion (Cl−) ausgenommen, so steht ein Hydrogencarbonatanion (HCO3−) als Austauschion zur Verfügung (siehe auch Abschnitt 17.2.2).
Zusätzlich kann über Protonenpumpen in der Zellwurzelmembran (H+) nach außen geliefert werden. Es bleibt Ladungsneutralität gewahrt!
Die Aufnahme von Ionen außen in die Wurzeln geschieht aber normalerweise gegen einen Konzentrationsgradienten und braucht daher Energie. Beim Transportmodell nach Van Den Honert und Epstein werden die Ionen selektiv mit Trägerproteinen transportiert. Über Hydrolyse stellt ATP (Adenosintriphosphat) dazu die Energie bereit.
Die Nährsalzelemente liegen im Substrat in vier Zustandsformen vor:
|
Bodenprozesse wie Nitrifizierung werden im Abschnitt 17.5.1 behandelt.
| Etwa 98 % des Bodens besteht aus schwerlöslichen Mineralien (Sulfate, Phosphate, Carbonate), Humus und anderem org. Material, welches nur langsam durch Verwitterung freigesetzt wird. Ungefähr 2 % der Düngerelemente sind an geladene kolloide Bodenteilchen (Ionen-Austauscher) gebunden. Und ein verschwindender Anteil von ungefähr 0,2 % sind als im Wasser gelöste und frei bewegliche Nährelemente vorhanden. |
Bringt man Ammoniumsulfat, wie auch andere Salze in den Boden ein, so wird es durch das vorhandene Wasser in Lösung gehen. Natürlich nur solange, bis eine Sättigung im Wasser eintritt. Ammoniumsulfat dissoziiert beim Lösen in Wasser in 2 Ammoniumionen und 1 Sulfation. Hier findet der reversible Zerfall einer Verbindung in Anionen und Kationen in einem Lösemittel statt, man spricht auch von elektrolytischer Dissoziation in einem Lösungsmittel. Es kommt zu einer Teilung einer chemischen Verbindung. In diesem Fall wird ein Molekül geteilt:
|
Schwefel nimmt die Pflanze ebenfalls aus dem Boden in Form von Sulfat (SO4) auf. Dieser wird auch zur Aminosäuresynthese benötigt.
So dissoziiert das Düngemittel Ammoniumnitrat NH4NO3 im Wasser folgendermaßen:
|
Wobei sich ein Dissoziationsgleichgewicht zwischen NH4OH ⇌ NH4+ + OH− einstellt. Der gesamt pH-Wert ist etwas sauer. Der pH-Wert von Ammoniumnitrat in wässriger Lösung bei einer Konzentration von 0,1 g/l und 25 ∘C stellt sich auf 6,1 ein. Eine Konzentration von 1 g/l ergibt pH-Wert von 5,6.
Die Wurzel kann Nährsalze letztendlich nur als im Wasser gelöste Ionen aufnehmen. Aus diesem Grund sollte der Boden ausreichend feucht sein, um die Aufnahme dieser zu ermöglichen!
Bei Huminstoffen sind die negativen Ladungen durch COOH- und phenolische OH-Gruppen bedingt, von denen mit steigendem pH-Wert Protonen (H+) abgespalten werden. In extrem saurem Milieu (pH-Wert < 3) können die organischen Bodenkolloide infolge einer Protonenanlagerung zu Anionentauschern werden. Sie besitzen daher ebenso wie Oxide und Oxidhydroxide eine variable Ladung.
Zwischen aufweitbaren (quellfähigen) Tonmineralien (Schichtsilikaten) können Wasser und Nährstoffe eingelagert werden und können als Ionenaustauscher für Pflanzen agieren.
Zweischichttonmineralien wie Kaolinit (Porzellanerde) sind in einer Schicht ein SiO4-Tetraeder-Gitter, wobei in jeder Ecke ein Sauerstoffion sitzt. Eine zweite Schicht ist als AlO6-Oktaeder organisiert. Diese sind kaum quellfähig und daher als Ionenaustauscher ungeeignet.
Bei Dreischichttonmineralien wie Montmorillonit, Illit, Smectit oder Vermiculit ist die Oktaeder-Schicht von zwei Tetraeder-Schichten begrenzt.
Der Aufbau der Silikate bei Dreischichtmineralien ist eine mittlere Schicht von MeO6-Oktaedern. Wobei Me als Platzhalter für Aluminium (Al3+), Eisen (Fe2/3+) oder Mangan (Mn2+) steht. Und zwei 2 Deckschichten aus SiO4-Tetraedern. Die Gesamtladung dieses Gitters ist negativ.
Zum Ladungsausgleich werden reversibel Kationen wie Na+, Ca2+ angelagert. Die negative Ladung ist unabhängig vom pH-Wert der Bodenlösung und sind somit Kationentauscher.
Vierschichttonmineralen wie die Chlorite sind nicht aufweitbar und daher als Ionenaustauscher ungeeignet. Diese entstehen in sauren Böden aus
Dreischichtsilikaten.
Enthalten Böden einen hohen Anteil feuchter Tonmineralen und Huminstoffen und liegt der pH-Werte der Bodenlösung zwischen 3 und 8, dann ist die Ladung der Tonmineralien negativ und es überwiegt der Kationenaustausch stark gegenüber dem Anionenaustausch ??.
Bei einem Ionenaustausch werden niederwertige Ionen von höherwertigen wegen der stärkeren elektrostatischen Anziehungskräfte vom Austauscher verdrängt. Al3+ verdrängt Ca2+ und Ca2+ verdrängt Na+. Beim Anionentausch verdrängt z.B. Phosphat PO43− Sulfat SO42−.
Bei gleicher Wertigkeit verdrängt das Ion mit der größeren Masse (molaren Masse) das Ion mit der niedrigeren. So verdrängt beispielsweise K+ (39 g/mol) das Na+ (23 g/mol).
Die austauschbaren Kationen sind von einer Hydrathülle umgeben. Unter gleichwertigen Kationen ist der Durchmesser der hydratisierten Ionen entscheidend (Ionengröße). Je dünner die Hydrathülle, umso besser ist die Anlagerung an den Austauscher: Mg2+ > Ca2+ und Na+ > K+. Die Art der Tonminerale spielt beim Austausch eine Rolle – so bevorzugt beispielsweise Montmorillonit Na+, K+. Berücksichtigt man lediglich die Ioneneigenschaften, so ergibt sich hinsichtlich Eintauschstärke und Haftfestigkeit meist nach der Hofmeister Reihe:
Es gibt aber auch eine Selektivität für Kalium K+, Ammonium NH4+-Ionen in illitischen Tonmineralen, die bevorzugt adsorbiert werden und die den oben angeführten Regeln nicht entsprechen.
Wenn der Boden saurer wird (der pH-Wert sinkt), werden mehr Protonen (H+) an die Bodenkolloide gebunden. Andere Kationen werden von den Bodenkolloiden ins Bodenwasser gedrängt. Steigt der pH-Wert, so werden die verfügbaren Kationen im Bodenwasser verringert und wieder an die Bodenkolloide gebunden.
Ein ganz niedriger pH-Wert kann bei tonigen Böden toxische Konzentrationen von Aluminiumkationen Al3+ freisetzen! Die Tonmineralzerstörung beginnt bei etwa einem pH-Wert von 4,5 und damit zu einer Freisetzung von Metallkationen in der Reihenfolge Mangan, Aluminium und Eisen.
Das dominierende Kation, mit dem K+ in Bodenkolloiden konkurriert, ist Calcium (Ca2+). Genauer nach [WK]: 60–90 % Calcium (Ca), 5–15 % Magnesium (Mg), 2–5 %, Kalium (K), 0–1 % Natrium (Na)
Die Bestimmung der austauschbaren Kationen erfolgt in Österreich nach ÖNORM L1086.
Solange Ammonium NH4+-Ionen adsorbiert sind, wird es nicht nitrifiziert. Bei der Adsorption an Tonmineralien verhält sich das Ammoniumion sehr ähnlich wie das Kaliumion, da sie gleiche Ladung und etwa gleichen Durchmesser besitzen ( K+ = 2,66 Å, NH4+ = 2,86 Å ). Daher kommt es bei hohen Kaliumgaben auch zu einer höheren Verfügbarkeit von Stickstoff, da Ammoniumionen in Lösung gehen.
Bei stark sauren Böden sind die Bodenkolloide von Protonen/Wasserstoffkationen (H+) besetzt und es besteht daher immer die latente Gefahr der Nitratauswaschung (NO3−) in das Grundwasser wie z.B. bei übermäßiger Düngung mit Gülle.
Bei stark verwitterten Böden in den Tropen mit viel Fe-Oxid oder Al-Oxid können sich die Verhältnisse umkehren. Hier sind vor allem unter sauren Bedingungen mehr positive Ladungen im Substrat vorhanden und daher können mehr Anionen gebunden werden. Die Konzentration austauschbarer Kationen ist sehr gering.
Die Summe aller austauschbaren Kationen eines Bodens ist die Kationenaustauschkapazität (KAK). Je höher der Anteil organischer Austauscher im Boden und je höher der pH-Wert des Bodens ist, desto größer ist die verfügbare/effektive KAK. In sauren Böden (pH-Wert < 7) ist die effektive KAK geringer, da hier angelagerte Wasserstoffionen (H+) Austauscherplätze zum Nachteil von anderen Kationen belegen.
| Böden mit hoher Ionenaustauschkapazität (KAK) binden auch Wasserstoffionen, deshalb verschiebt sich der optimale Pflanzen-pH-Wert nach oben ??. |
In carbonathaltigen Böden sind an der KAK fast ausschließlich Ca2+, Mg2+, K+ und Na+-Ionen beteiligt.
|
In basischen bis sauren humusreichen oder tonigen Böden lagern sich Ca2+, Mg2+, K+, Na+, Al3+, NH4+-Kationen an den
Ton bzw. Humus an. Durch Abgabe von Protonen/Wasserstoffkationen (H+) von Wurzel, werden schwach gebundenen Kationen (pos. geladene Ionen) gelöst
und dann von den Wurzeln aufgenommen – Ionenaustausch. Dabei bleibt die Ladungsneutralität erhalten. Auch Anionen werden, wenn auch in weit geringerer Menge, wie gebundene PO43−, SO42−, NO3−, Cl−-Anionen gegen Hydrogencarbonatanionen (HCO3−) der Wurzeln ausgetauscht. |
Gegenspieler (Antagonismen) und Synergismen bei der Nährstoffaufnahme: Die einzelnen Nährstoffe stehen in gegenseitigen Wechselbeziehungen. Je nach Zusammensetzung der verschiedenen Düngernährstoffe in der Bodenlösung, kann eine Konkurrenzsituation auftreten. Ein Übermaß des einen Nährstoffs blockiert die Aufnahme eines anderen. Aber auch das Gegenteil ist möglich: gewisse Nährstoffe fördern die Aufnahme gegenseitig. Diese Wechselwirkungen sind teils pflanzenabhängig!
Die folgende Teilauszug aus [Hau12] mit Anpassungen zeigt die wichtigsten dieser Wechselbeziehungen.
| hohe Konzentration von | erschwert die Aufnahme von | fördert die Aufnahme von |
| NH4 (Ammonium) | Ca, Mg, K | P, SO4 (Sulfat) |
| NO3 (Nitrat) | PO4 (Phosphat) | Ca Mg, Mn, K |
| Ca (Calcium) hemmt auch KAK | Mg, K, Fe, B, Mn | |
| K (Kalium) | Ca, Mg, NH4 | NO3 |
| Mg (Magnesium) | Ca, K, NH4 | |
| P (Phosphor) | Fe (Ca, B,Cu) | Zn |
| Na (Natrium) | Ca | P |
| Cu (Kupfer | Fe | P |
| In Europa wird der Gehalt von Phosphor, Kalium und Magnesium auf Düngern nicht direkt angegeben, sondern es werden für die Oxid-Formen (Phosphorpentoxid P2O5, Kaliumoxid K2O und Magnesiumoxid MgO) die Massenprozente angegeben. |
Ein (20/10/8+2) NPK+Mg-Dünger enthält in äquivalenter Form 20 N, 10 P2O5, 8 K2O + 2 MgO %.
Umrechnungsfaktoren für die Massen reiner Elemente:
| Phosphor (P) = | Phosphorpentoxid (P2O5) | x 0,436 |
| Kalium (K) = | Kaliumoxid (K2O) | x 0,830 |
| Magnesium (M) = | Magnesiumoxid (MgO) | x 0,603 |
Der Stickstoff wird in der Pflanze vor allem für das Wachstum gebraucht; man bezeichnet ihn auch als „Motor des Wachstums„, weil er als Bauelement sowohl in Chlorophyll, Enzymen und Eiweißen enthalten und somit unter anderem für das Blätter- und Längenwachstum entscheidend ist.
Aufgenommen wird Stickstoff generell über die Wurzeln, in Form von Nitrat-Anionen (NO3−) und als Ammoniumkationen (NH4+). Pflanzen, die aufgrund der Durchlässigkeit der Wurzelmembranen vorwiegend NH4+ aufnehmen, sind auf saure Böden angewiesen. Daraus kann man ableiten, dass Pflanzen, die gut auf basischen Böden gedeihen, mehr Nitratanionen als Ammoniumkationen aufnehmen. Umgekehrt nehmen Pflanzen mehr Ammonium auf, die gerne auf sauren Böden wachsen.
Geht man von der Stickstoffstudie [PT03] aus und nimmt an, dass sich die Eiche (Quercus rubra) bei der Stickstoffaufnahme ähnlich der Edelkastanie verhält, so kann das Massenaufnahmeverhältnis bei etwa
| Ammonium (NH4+) : Nitrat ( NO3−) = 2,5 : 1 |
liegen.
Der Vorteil des Nitrats ist die freie Beweglichkeit der Ionen im Boden. Die Nitrationen sind im Bodenwasser gelöst und
gelangen schnell und leicht an die Wurzeln.
| Die drei Hauptstickstoffquellen in der Landwirtschaft sind Ammonium-Verbindungen, Nitrat-Verbindungen und Harnstoff. |
Da viele Pflanzen nur zum Teil Ammonium-Verbindungen direkt aufnehmen, ist die Umwandlung von Stickstoffverbindungen durch enzymatische Reaktionen in Verbindung
von Bakterien ein wesentlicher Bestandteil zum Pflanzenwachstum:
Urease von Harnstoff
|
Ammonifizierung
|
Nitrifizierung
|
Nitrifizierung in vollständiger Ionenform ohne Ammonium
|
| Die beiden verbleibenden 2 Wasserstoffkationen senken den pH-Wert im Boden. |
Harnstoff (CH4N2O : H2N−CO−NH2 ) ist das Endprodukt des Eiweißstoffwechsels bei Tier und Mensch. Er wird in der Leber aus
Ammoniak (NH3) und Hydrogencarbonaten (HCO3−) gebildet. Täglich scheiden die Nieren eines Menschen
etwa 13–33 g Harnstoff aus.
Harnstoff wird im Boden über den Prozess der Urease zunächst in Ammoniak umgewandelt.
Durch Urease-positive Bakterien über das Enzym Urease wird innerhalb von 1 bis 4 Tagen, der eingebrachte
Harnstoff plus Wasser, in Ammoniak und Kohlendioxid gewandelt.
Mischt man das Gas Ammoniak in Wasser so werden sich zum Teil Nitratkationen (NH4+) und Hydroxidanionen (OH−)
als alkalisches Ion bilden. Es stellt sich je nach Temperatur ein Gleichgewichtszustand ein – Ammonifizierung.
Je höher die Temperatur ist, desto höher wird der Ammoniakanteil (NH3) sein und der pH-Wert sinkt.
Ist die Umgebung sauer, dann geht der Ammoniak-Anteil zurück, unter pH-Wert von 6 gibt es keinen Ammoniak im Wasser.
Löst man eine Ammoniumverbindung (NH4+) in Wasser, so wird sich auch Ammoniak laut obiger Gleichung bilden.
In der ersten Phase der Nitrifzierung wird mit Hilfe von Nitritbakterien (Nitrosomonas) durch eine enzymatische Reaktion und molekularen Sauerstoff, Ammonium (NH4+) zu salpetriger Säure (HNO2), einer Nitritverbindung (NO2− ), plus Wasser und 2 Wasserstoffionen oxidiert.
In einem zweiten Schritt wird durch Nitratbakterien (Nitrobacter) die salpetrige Säure bzw. Nitrit (NO2−) durch enzymatische Reaktion mit Sauerstoff zu Nitrat (NO3−) oxidiert. Bei beiden Prozessen wird Energie freigesetzt. Der gesamte Prozess der Nitrifizierung dauert je nach Bodentemperatur 2 bis 6 Wochen. Die beiden verbleibenden 4 H+ Wasserstoffionen, wobei sich zwei wieder mit OH zu Wasser vereinen, senken den pH-Wert im Boden.
Um das Nitrat den Pflanzen nicht auf einmal zuzuführen, kann man den Nitrifizierungsprozess hinauszuzögern – dazu verwendet man sogenannte Nitrifikationshemmer. Diese vermindern die Aktivität von Nitrosomonas und Nitrobacter für etwa vier bis zehn Wochen (z.B. Dimethylpyrazolphosphat C5H11N2O4 P und Dicyandiamid C2H4N4).
|
Für Urease liegt der optimale pH-Wert in einem Bereich 6,5–8,0; wobei der Wert bei höherer Harnstoffkonzentration noch oben wandert.
Die Ureaseaktivität durch Bakterien soll in einem Bereich von 2 ∘C bis 45 ∘C linear ansteigen. Das Optimum der Bodentemperatur für die Nitrifizierung liegt bei 26∘C. Bei 12 ∘C wird der Prozess deutlich verlangsamt und bei 5 ∘C findet keine Nitrifizierung statt und bei 0 ∘C sterben die Bakterien. Optimale Feuchte ist 60 % der maximalen Wasserhaltekapazität. Oberhalb von 90 % wirkt der Sauerstoffmangel limitierend. Der optimale pH-Wert liegt bei etwa 7,8–8,0 für Nitromonas und 7,3–7,5 für Nitrobacter. Unter optimalen Bedingungen verdoppelt sich die Anzahl der Nitrosomonas alle 7 Stunden und Nitrobacter alle 13 Stunden was für Bakterien relativ langsam ist. Das Wachstum der Nitrosomonas wird bei pH-Wert von 6,5 unterbunden. Bei einem pH-Wert von 6,0 wird die Nitrifizierung eingestellt. [Lab09]. Da in sauren Waldböden Ammonium (NH4+) nicht so intensiv nitrifiziert wird, haben Bäume sich daran angepasst und nehmen Ammonium im großen Maße direkt auf. |
Nach der Urease kann der gasförmige Ammoniak (NH3) nur zum Teil vom feuchten Boden gehalten werden – Ammonifizierung. Und wird, besonders bei trockenen Böden, auch als Schadstoff in die Luft emittiert.
Weiters sind Urease-positive Bakterien in den Exkrementen vorab die Ursache für unerwünschte Ammoniak-Emissionen in der landwirtschaftlichen Tierhaltung und gehen schon bei der Lagerung oder oberflächlichen Ausbringung von Jauche verloren. Daher sollte Jauche unter Luftabschluss gelagert werden.
Bei einem Mangel an Stickstoff in der Pflanze kann auch zu einer Chlorose (Chlorophyllmangel) kommen, bei der die Blätter gleichmäßig hell- bis gelbgrün werden, zum anderen kann eine Nekrose (= Absterben) der älteren Blätter durch Eiweißabbau entstehen oder der Mangel macht sich durch ein schwaches Wachstum aufgrund einer Störung des Stoffwechsels bemerkbar.
Nitrat wird im Boden fast nicht gebunden und gelangt bei einer Überdüngung schnell ins Grundwasser. Eine Überdüngung der Pflanze mit Stickstoffverbindungen führt auch in der Pflanze selbst zu einer erhöhten Nitratkonzentration. Nitrat kann im Körper zu Nitrit und schließlich zu Nitrosaminen umgewandelt werden, die krebserregend sind.
Bei einem Mangel an Eisen in der Pflanze kann es zu einer sogenannten Christbaum-Chlorose kommen, bei der die Blätter geblich sind, während die Blattadern grün sind. Bleibt der Eisenmangel bei wachsenden Blättern bestehen, vertrocknen die Blätter später vom Rand her. Anmerkung: Junge Blätter, besonders nach dem Erstaustrieb im Frühjahr bleiben für einige Zeit etwas geblich und auch etwas rötlich, was sich später wieder gibt!
Der Eisenmangel ist meist kein primärer Eisenmangel, sondern die Aufnahme von Eisen wird durch kalkhältige Böden gehemmt. In der Regel ist das Eisen ein Bestandteil von Eisenoxiden und damit nur schwer löslich, so dass es von den Wurzeln der Pflanzen nicht unmittelbar absorbiert werden kann. Ein höherer Säuregehalt – niedriger pH-Wert – im Boden löst die Eisenoxide an und macht so zweiwertige Eisenionen (Fe2+) an den Wurzeln verfügbar. Anmerkung: Pflanzen senken den pH-Wert im Wurzelbereich auch selbst (siehe auch 17.2.2) um Nährstoffe besser aufnehmen zu können.
Abbildung 110: Chlorose: nach Austrieb li, fortgeschritten re
Bei einem profunden Eisenmangel werden gelbe Blätter auch bei nachträglicher Behebung des Eisenmangels nicht mehr richtig grün und unterstützen die Pflanze bei der Photosyntese nur mehr gering.
Eine einfache aber etwas teure Möglichkeit zur Behebung der Problems, auch bei kalkhältigen Böden, ist eine Gießwasserdüngung mit Eisenchelat Fe-EDDHA – geeignet für pH-Werte von 2,5 bis 11,0. Dieses wird z.B. als Sequestrene 138 Fe (6 % Fe-wasserlösliches Eisen) von Syngenta hergestellt. Kwizda, Compo und andere vertreiben diesen Eisendünger.
Das Eisenchelat Fe-EDTA ist ungeeignet, da es im Boden nur bis pH-Wert 6 wirksam ist. Tritt der Chlorosemangel jährlich auf, so sollte man schon beim Knospenaustrieb Fe-EDDHA übers Gießwasser in den Boden einbringen. Eine Düngung mit Fe-EDDHA zwei bis dreimal im Jahr in der Wachstumsphase ist meist ausreichend. Die Gießwassermischung ist möglichst rasch zu verbrauchen, da das aufgelöste Chelat nicht lange stabil bleibt.
Eine Senkung des pH-Werts des Bodens, wie im Abschnitt 18.2.3 gezeigt, ist nur mit großem Aufwand möglich.
| Zusammenfassung: Nitrathaltige Düngemittel sind für Pflanzen schnell verfügbar. Ammoniumverbindungen können von Waldbäumen auch direkt aufgenommen werden. Harnstoffe und auch Ammoniumverbindungen werden über mehrere Schritte im Boden mit Hilfe von Bodenbakterien, Sauerstoff und Wasser in Nitrate oxidiert. |
Phosphat ist in der Pflanze an allen wichtigen Stoffwechselprozessen wie Blüten-, Frucht- und Samenbildung beteiligt und daher ein wichtiger Energieträger. Darüber hinaus stellt es einen Baustein der Zellmembran dar.
Phosphat unterliegt im Boden der Alterung, d. h. gedüngtes Phosphat wird im Boden recht schnell adsorbiert und steht den Pflanzen fortan nur eingeschränkt zur Verfügung. Eine Phosphatdüngung sollte also möglichst zeitnah zum Bedarf der Pflanze ausgebracht werden, um kurzfristig eine Erhöhung der Phosphatkonzentration im Boden zu erreichen.
Bei einem Phosphatmangel kommt es aufgrund von Stoffwechselstörungen zu einer rötlichen Blattfärbung (Chlorophyll- und Anthocyan–nhäufung), und zu einer schlechten Gesamtentwicklung in Bezug auf die Blätter, Wurzeln und Blüten. Bei einer Überdüngung und somit bei erhöhtem Phosphatgehalt, kann es in Gewässern zu einem vermehrten Auftreten von Wasserpflanzen und Algen kommen.
Kalium ist vor allem für die Steuerung des Wasserhaushalts in der Pflanze verantwortlich. Die Kaliumkonzentration regelt den osmotischen Druck in den Zellen und dadurch das Öffnen und Schließen der Spaltöffnungen. Darüber hinaus fördert es die Fotosyntheseaktivität und damit auch die Bildung von Stärke bzw. Zucker.
Bei Kaliummangel kommt es vor allem bei älteren Blättern zu Blattrandnekrosen und auch zu einer erhöhten
Krankheitsanfälligkeit der Pflanze [SF09].
Bei den organischen Düngern sind, oder waren, die düngenden Elemente meist in kohlenstoffhaltigen reduzierten Verbindungen gebunden. Sind diese bereits, wie etwa im Kompost, teilweise oxidiert, so sind die düngenden Mineralien an den Abbauprodukten (Huminsäuren) etc. adsorbiert [Wik15c].
| Pflanzen können direkt keine organische Verbindungen aufnehmen, diese müssen erst durch Zersetzung mineralisiert werden. |
Gülle ist eine Mischung aus Kot und Harn der landwirtschaftlichen Nutztiere Schwein und Rind. Mist ist fest und besteht aus Urin, Kot und aus einem Bindemittel, üblicherweise Stroh. Sickersäfte aus dem Misthaufen bezeichnet man als Jauche. Sie besteht hauptsächlich aus vergorenem Urin und Regenwasser, wenn ein Misthaufen unter freiem Himmel gelagert wird.
Der Harnstoff im Urin wird durch Urease schon vor der Ausbringung in das Ammoniak (NH3) und Kohlendioxid (CO2) zerlegt. Luftzutritt fördert die Harnstoffumwandlung stark und damit die Verflüchtigung von Stickstoff über Ammoniak. Harn ist reich an Stickstoff und Kali, enthält aber nur wenig Phosphor.
Auszug der mittleren Nährstoffgehalte organischer Dünger[NW14] in gerundeten Werten an der Frischmasse pro tonne (t):
| TM % | Gesamt −N | P2O5 | K2O | MgO | CaO | |
| Rindermist | 23 | 6 | 3 | 10 | 2 | |
| Schweinemist | 23 | 7 | 7 | 7 | 3 | |
| Pferdemist | 32 | 5 | 3 | 10 | 2 | |
| Putenmist | 50 | 19 | 18 | 16 | 6 | 20 |
| Hühnerfrischkot | 30 | 17 | 11 | 10 | 8 | 50 |
| Grünschnittkompost | 61 | 7 | 3 | 6 | 5 | 17 |
TM bezeichnet den Anteil an Trockenmasse in % an der Gesamtfrischmasse. Kann auch als g pro 1 kg an der Gesamtfrischmasse (Gesamtfeuchtmasse) gesehen werden.
Für Rindermist bedeutet das 6 kg Gesamtstickstoff (N) pro Tonne (t) Mist. Damit ergeben sich (0,6/0,2/0,3) % NPK-Düngeranteile zur Gesamtmasse. Für die Anteile an der theoretischen Trockenmasse (0 % Wasser) ergeben sich (2,6/0,86/1,3) %.
Für einen Hühnertrockenkot ergeben sich die (5,6/3,6/3,3) % NPK-Düngeranteile.
Der früher in großen Mengen abgebaute Vogel-Guano – exemplarisch hier angeführt der Peru Guano (nach Knickmann), enthält etwa (8-15/2-5/2-4) % NPK-Düngeranteile zur Gesamtmasse[KS16].
Abgepackte organische Zukaufdünger werden meist aus pelletiertem Rindermist, Pferdemist, Hühnermist, Hornspänen, Knochenmehl, Kompost, Ernterückständen wie Traubentrester, Melasse, Mälzereirückstände und Pilzmycel aus der Vitamin C und der Penicillinerzeugung (Handelsprodukte wie Biosol, Biofert) angeboten.
Pilzmycel enthält einen hohen Anteil von Chitin, welches angeblich gegen pathogene Pilze wie z. B. Pyrenochaeta lycopers hilft.
Organische Dünger pflanzlichen Ursprungs haben den Vorteil, dass die mineralisierten Nährstoffe langsam und weitgehend gleichmäßig verfügbar sind. Dadurch wird das
Feinwurzelsystem der Pflanzen nicht angegriffen und Mykorrhiza können sich gut ausbilden.
Unter dem Markennamen Biosol wird ein Abfallprodukt der Penicillinproduktion
als organischer Dünger angeboten. Mit Hilfe von Ausgangsstoffen wie Sojamehl, Baumwollsamenmehl, Zucker, Sirup und Vitaminen
wird über einen Fermentationsprozess mit Hilfe des
Schimmelpilzes Penicillium chrysogenum, Penicilline produziert. Aus der Fermentationsbrühe werden die Antibiotikawirkstoffe isoliert und
das anfallende Pilzmycel bei 110−130 ∘C für 3–4 Stunden getrocknet, damit auch sterilisiert und zu Dünger verarbeitet [Nas95].
Biosol Inhaltsstoffe [San15] (2015):
| Organische Substanz | 85 % | |
| Stickstoff org. gebunden | 6-8 % | |
| Stickstoff wasserlösl. max. | 0,5 % | |
| Phosphat (P205) | 0,5 % | |
| Kaliumoxid (K20) | 0,3 % | |
| C/N Verhältnis | 6:1 |
Biosol Forte Inhaltsstoffe (2015):
| Organische Substanz | 70 % | |
| Stickstoff org. gebunden | 6-9 % | |
| Phosphat (P205) | 3 % | |
| Kaliumoxid (K20) | 0,5 % | |
| C/N Verhältnis | 6:1 |
Unter dem Markennamen Biofert wird ein Abfallprodukt der Zitronensäureproduktion als
organischer Dünger angeboten.
Biofert Inhaltsstoffe [KG15] (2015):
| Organische Substanz | min. 40 % | |
| Stickstoff gesamt | 4 % | |
| Phosphat (P205) | 1,5 % | |
| Kaliumoxid (K20) | 0,5 % | |
| Calciumoxid (Ca0) | 11 % | |
| Zink (Zn) | 0.05–0,25 % |
Allgemein wird für die Zitronensäureproduktion mit den Ausgangsstoffen wie Zucker, Melasse, Glucosesirup über einen Fermentationsprozess mit Hilfe von Aspergillus niger-Schimmelpilzen Zitronensäure produziert. Achtung: Biofert enthält viel Calcium.
Im anorganischen Dünger oder Mineraldünger sind die Düngeelemente meist in Form von Salzen gebunden:
Stickstoff wird meist in Form von Ammoniumnitrat (H4N2O3), Ammoniumsulfat ((NH4)2SO4), Kaliumnitrat (KNO3−), Harnstoff (CH4N2O) auch Carbamidstickstoff genannt oder als Formaldehydharnstoff (hohe Lagerstabilität) in den Boden eingebracht.
Phosphate werden in z.B. als Diammoniumphosphat ((NH4)2HPO4) – Monoammoniumphosphat (MAP) gebunden in Dünger angeboten. Im Bergbau gewonnenes Phosphorit, ein marines Sedimentgestein, welches ein Gemenge aus Calcium-Fluorapatit (F), Hydroxylapatit (OH): Ca5(PO4)3 [F,OH,CO3] und organischen Substanzen ist, muss erst mit Schwefelsäure (H2SO4) in wasserlösliches Calciumdihydrogenphosphat (Ca(PO4)2 ) + wasserunl. Calciumsulfat (CaSO4), auch Superphosphat genannt, aufgeschlossen werden.
Kalisalze sind ein Gemisch aus verschiedenen Salzen (NaCl, KCl, KMgCl3, MgSO4), diese werden im Bergbau gewonnen,
dann aufbereitet und in Kaliumchlorid (KCl) oder zu Kaliumsulfat (K2SO4) umgesetzt.
In [Smi11] und [Sch11] wird auf eine mögliche Schwermetallbelastung in anorganischen Düngern hingewiesen.
NPK-Dünger Kennzeichnung
| (13/13/21) | rot gefärbt oder gekennzeichnet, Kalium in Chloridform |
| (15/15/15) | gelb gefärbt oder bezeichnet, Kalium in Chloridform |
| (12/12/17 + 2) | blau gefärbt, bekannt unter Markennamen Blaukorn, Kalium als Sulfat, 2 % MgO. |
Immergrün Blaukorn (15/6/12/+2)
| 15 % | N | Gesamtstickstoff | |
| 13 % Ammoniumstickstoff | |||
| 2 % Carbamidstickstoff | |||
| 6 % | P2O5 | neutral-ammoncitratlösliches und wasserlösliches Phosphat | |
| 5,0 % P2O5 wasserlösliches Phosphat | |||
| 12 % | K2O | wasserlösliches Kaliumoxid | |
| 2 % | MgO | 1,6 % MgO wasserlösliches MgO | |
| 0,02 % B Bor; 0,06 % Fe Eisen; 0,01 % Zn Zink |
COMPO Rhododendron Langzeit-Dünger (16/5,5/14/+2)
| 16 % | N | Gesamtstickstoff | |
| 6,8 % Nitratstickstoff – Ammoniumnitrat | |||
| 9,2 % Ammoniumstickstoff | |||
| 5,8 % | P2O5 | neutral-ammoncitratlösliches und wasserlösliches Phosphat | |
| 4,9 % P2O5 wasserlösliches Phosphat | |||
| 14 % | K2O | wasserlösliches Kaliumoxid | |
| 2 % | MgO | 1,4 % MgO wasserlösliches MgO | |
| 0,02 % B Bor; 0,02 % Cu Kupfer; 0,2 % Fe Eisen;Mn Mangan; 0,006 % Zn Zink |
Dehner Blütenzauber – wasserlöslich (15/10/10+2/13) 1kg
| 15 % | N | Gesamtstickstoff | |
| 3 % Nitratstickstoff | |||
| 12 % Ammmoniumstickstoff | |||
| 10 % | P2O5 | wasserlösliches Phosphat | |
| 10 % | K2O | wasserlösliches Kaliumoxid | |
| 2 % | MgO | 1,6 % MgO wasserlösliches MgO | |
| 13 % | S | wasserlöslicher Schwefel | |
| 0,025 % B Bor; 0,01 % Cu Kupfer; 0,1 % Fe; 0,05 % Mn Mangan; | |||
| 0,002 % Mo Molybdän; 0,025 % Zn | |||
| Cu, Fe, Mn, Zn als Chelat von EDTA |
COMPO Blumenprofi – wasserlöslich (11/14/17+2) 1,2kg
| 11 % | N | Gesamtstickstoff | |
| 1,5 % Nitratstickstoff | |||
| 9,5 % Ammmoniumstickstoff | |||
| 14 % | P2O5 | wasserlösliches Phosphat | |
| 17 % | K2O | wasserlösliches Kaliumoxid | |
| 2 % | MgO | 1,6 % MgO wasserlösliches MgO | |
| 0,03 % B Bor; 0,16 % Cu Kupfer; 0,11 % Fe; 0,16 % Mn Mangan; | |||
| 0,023 % Mo Molybdän; 0,06 % Zn | |||
| Cu, Fe, Mn, Zn als Chelat von EDTA |
Dünger, Gießwasser, Mikroorganismen und Pflanzen verändern den pH-Wert. Das Substrat selbst unterliegt auch einer zeitlichen Änderung.
Der pH-Wert des Substrates beeinflusst die Verfügbarkeit von Nährsalzen für die Pflanze. Außerdem ist der pH-Wert meist ein Indikator für den Calciumkarbonatgehalt des Bodens bzw. Substrates.
Ein ausführliches und gut aufbereitetes Skriptum über Bodenkunde der Professur für Bodenökologie der Universität Freiburg findet sich unter [Fre99].
Die Gesamthärte gibt die Summe der Konzentrationen der Kationen von Erdalkalimetallen im Wasser an. Die Carbonathärte ist derjenige Anteil der Gesamthärte, der alkalisch wirkt und damit den pH-Wert des Substrats beeinflusst. Die Härte des Wassers wird durch bestimmte Calcium- und Magnesiumsalze verursacht.
Die Gesamthärte wird gegliedert in die:
Carbonathärte (vorübergehende Härte) – Menge an Calciumcarbonat/kohlensaurer Kalk (CaCO3) und Magnesiumcarbonat (MgCO3)
Nichtcarbonathärte (bleibende Härte) – Menge an Calciumsulfaten und Magnesiumsulfaten bzw. -chloriden
Die Carbonathärte, wird aus dem Hydrogencarbonat (HCO3− = OH CO2−) gebildet, die durch Auflösung
von kohlensaurem Kalk in Wasser und Kohlensäure entstehen. Nur diese ist ausschlaggebend für den pH-Wert.
|
Anmerkung: Oberhalb einer Temperatur von 50 ∘C zerfällt Hydrogencarbonat und die linke Seite des Reaktionsgleichgewichtes dominiert.
Kalkhaltiges, also hartes Gießwasser, erhöht den pH-Wert. Die zugehörige Einheit ist die deutsche Härte dH∘.
1∘dH entspricht 10mg CaO/l oder 17,8 mg CaCO3/l.
Folgende grobe Abstufung der Wasserhärte ist gebräuchlich:
|
In Österreich sind laut Bodenuntersuchungen über 20 % der Ackerböden und 60 % der Grünlandböden als sauer (pH-Wert unter 5,5) einzustufen [Gal13]. Über 45 % aller Grünland- bzw. 35 % aller Ackerböden sind als kalkfrei einzustufen.
Substrate bzw. Böden haben normalerweise die Eigenschaft, trotz Zufuhr von Säuren (H+) oder Basen (OH−), den pH-Wert im einem bestimmten Bereich
konstant zu halten. Säurepuffer können Wasserstoffionen H+ aufnehmen. Der
pH-Wert und die sogenannte Pufferkapazität ist stark von der Bodenart abhängig
[Bod14a],[Fre99].
Säurepufferung:
| Puffersystem | pH-Pufferbereich | Pufferkapazität | Anmerkung | ||
| Carbonate | 6,5 –8 | 300 kmol H+ pro 1 % | Carbonat | Anionenüberschuss (Phosphat-, Fe-Fällung) | |
| Kationenaustausch | 5,5–7 | 25 kmol H+ pro 1 % | Ton | Auswaschung von Ca, Mg, K | |
| Silikate | 4,2–5,5 | 8 kmol H+ pro 1 % | Silikat | Kationen-/Anionenverhältnis ausgeglichen | |
| Aluminium–Oxide, Hydroxide | 3,0–4,8 | 150 kmol H+ pro 1 % | Ton | starke bis vollständige Kationenauswaschung, toxisch | |
| Eisen–Oxide | < 3,0 | extremer Nährstoffmangel, Fe3+ und H+ vorherrschend | |||
|
300 kmol Wasserstoffionen H+ pro 1 % Carbonat bedeutet, dass je % Calciumcarbonat im Boden (bis in 10 cm Tiefe) etwa 300 kg Wasserstoffionen pro ha neutralisiert werden können, ohne den pH-Wert zu ändern [Gal13]. Bei Böden mit ausreichend Calzium- oder Magnesium-Carbonaten, wirkt vor allen das Carbonat-Puffersystem. Carbonate werden bei pH-Werten über 6,5 gebildet. Fehlen die Carbonate, so erfolgt die Pufferung mit wesentlich geringerer Pufferkraft über den Austausch an den Bodenkoloiden. Diese absorbieren vor allem basische Kationen Ca+- und Mg+-Ionen im Austausch gegen die sauren H+-Ionen in der Bodenlösung. Ist dieser Puffer erschöpft, kommt das Silikat-Puffersystem zum tragen, welches über die Verwitterung von Silikaten, Basen freigesetzt. Unter pH 4,2 kommt es zum Zerfall der Tonminerale, wobei gleichzeitig wurzeltoxische Aluminium-Ionen freigesetzt werden. |
Die Folgende Gleichungen sind in Ionenform angeschrieben.
Der Carbonatpuffer ist in Böden mit hohem Kalksteinanteil (Calciumcarbonat) vorherrschend.
Bei Säureeintrag in den Boden werden die Protonen/Wasserstoffionen (H+) zur Auflösung von Calciumkarbonat (CaCO3) verwendet. Ist das Calciumkarbonat im Boden fein verteilt, so geschieht dieser
Umwandelungsprozeß verhältnismäßig rasch. Bei einem hohen Anteil von Calciumkarbonat wird sich ein pH-Wert um die 7 einstellen.
Bei Aufkalkung des Bodens werden Wasserstoffionen (H+) als basisch wirkendes Hydrogencarbonat (HCO3) gebunden [Bod14b]
Mit schwachen Säuren reagiert Calciumcarbonat (Calzit) gezeigt hier in Ionenform zu:
|
Mit starken Säuren zu
|
Auch Magnesiumcarbonat ist ein Puffer. Der Reaktionsablauf ist analog wie beim Calciumcarbonat:
|
Auch Dolomit ist ein Puffer:
|
Der Kationenaustausch erfolgt an den Bodenkolloiden. Dabei werden die Bodenkolloide gebundenen Calcium- und Magnesium-Kationen gegen Protonen/Wasserstoffkationen (H+) ausgetauscht. Siehe auch Abschnitt 17.2.2.
Der Silikatpuffer allgemein:
|
Und exemplarisch durch hier nach Auflösung (Verwitterung) von Kalifeldspat mit Säuren gezeigt:
|
Der Aluminiumpuffer beginnt bei Zerfall der Tonminerale zu wirken, wenn Tonböden sehr sauer (pH-Wert unter 4,2) sind. Das freigesetzte Aluminium wirkt toxisch auf die Pflanzenwurzeln.
|
Ein erhöhter Aluminiumgehalt im Grundwasser kann ein Hinweis auf eine extreme Bodensituation sein!
Der Eisenpuffer:
|
Diese Situation kann z.B. in Bergbaugruben auftreten und ist hier nur von theoretischer Bedeutung.
Zur Bestimmung des Carbonatanteiles soll der Gewichtsverlust durch Ausgasung von (CO2) bei Säurezugabe in die Erde herangezogen werden. Man könnte auch die Ausgasung auffangen und das Volumen bestimmen.
Achtung bei Umgang mit Säuren immer Handschule und Schutzgläser tragen!
Mit einem Erdbohrer mindestens in 3 Proben in 20 cm und 3 Proben in 40 cm Tiefe vom Acker an verschiedenen Stellen entnehmen. Diese Proben zusammenmischen und wieder kompaktieren. Etwa 1/4 l kompaktierte Erde in ein sauberes Glas geben und verschließen.
Ich habe die Messung auf einen Waagenbereich auf 20 g (Auflösung 0,001 g) ausgelegt.
Etwa 1/4 l Erde in einer flachen Schale in der Mikrowelle trocknen. Die Erdkrümmel zerdrücken und die trockene Erde gut durchmischen. Etwa 8 g in einen kleinen Becher einwägen. Als nächsten Schritt die Trockenmasse mTROCK exakt messen. Dann etwa 2–4 g dest. Wasser zugeben und leicht schwenken. Etwa 4 g einer 5–10 %igen Salzsäure vorbereiten. Den Becher auf die Waage stellen und die verdünnte Säure zügig zugeben und gleich darauf die Masse messen mVOR. Danach so lange warten bis kein Schaumblubbern auftritt. Dieses Blubbern sieht man nach einiger Zeit nicht mehr, man hört ihn noch gut. Wenn keine weiteren Bläschen entstehen, die Messung mNACH durchführen.
Gemessen:
| mTROCK | 7,649 g |
| mVOR | 16,800 g |
| mNACH | 12,558 g |
Massenverlust mCO2 = mVOR − mNACH = 12,680 − 12,558 = 0,124 g Entspricht einer Calciumcarbonat-Masse von mCaCO3 · 2,2742 = 0,186 · 2,2742 = 0,423 g
Anteil an Calciumcarbonat ACaCO3 = mCaCO3 / mTROCK · 100 = 0,423 / 7,649 · 100 ≈ 5,5 %
Der Trockenmassenateil (w/w) ist 5,5 % Carbonate.
Ein kalkhältiger Ackerboden soll für säureliebende Pflanzen adaptiert werden. Auf einer Fläche von 10x10 m bis bis in eine Tiefe von 0,5 m soll die Bufferkapazität der Carbonate chemisch neutralisiert werden. Es soll berechnet werden wie viel Schwefel oder Eisen-II-Sulfat in den Boden eingearbeitet werden muss.
Den pH-Wert langsam senken durch Ausschalten des Carbonatpuffers mit Schwefel zu Gips (innerhalb eines Jahres):
|
Thiobac Bakterien verarbeiten den Schwefel zu Schwefeltrioxid (SO3). Das Schwefeltrioxid (SO3) und Wasser reagieren zu Schwefelsäure (H2SO4).
|
Letztlich reagiert Schwefelsäure (H2SO4) mit Calciumkarbonat (CaCO3) zu Gips (CaSO4):
|
Anmerkung: Gips (CaSO4) hydradisiert mit Wasser ( 2H2O) in der Natur zu (Ca[SO4]· 2H2O).
Prüfen der linken und rechten Seite der Reaktionsgleichung:
|
Für die Wandlung von einem Mol Calciumcarbonat (CaCO3) mit 100,0869 g braucht man 32,06 g Schwefel (S).
|
|
Für die Wandlung von einem kg Calciumcarbonat (CaCO3) in Gips (CaSO4) braucht man 0,3098 kg Schwefel (S) und es werden
0,4397 kg Kohlendioxid (CO2) freigesetzt. Oder auch für die Fixierung von einem kg reinem Calciums (Ca) braucht man 0,7736 kg Schwefel (S)! Bei einem Carbonatanteil 5,5 % (großteils Calciumcarbonat) und einem spez. Trockengewicht der Erde von 1000 kg/m3 auf 100 m2 bis in eine Tiefe von 0,5 m errechnet sich der Carbonatanteil zu 0,055 · 100 · 0,5 · 1000 = 2750 kg . Man muss 2750 · 0,3098 kg ≈ 850 kg Schwefel in 100 m2 einarbeiten. Das wären 85 t/ha! |
Eisen-(II)-sulfat FeSO4 bindet die Luftfeuchtigkeit. Im Handel ist es normalerweise als Eisen(II)-sulfat-Heptahydrat FeSO4 7H2O mit blau-grüner Farbe zu finden.
|
und wieder mit der Schwefelsäure
|
| CaCO3 : FeSO4·7H2O : CaS04 : FeO : H2SO4 : H2O : Ges = 100,086 : 98,0785 : 136,1406 : 44,0095 : 18,0152 : 254,0136 g/mol |
|
|
|
Um einem kg Calciumbarbonat (CaCO3) in Gips (CaSO4) umzuwandeln braucht man 2,78 kg Eisen-(II)-sulfat-Heptahydrat (FeSO4·7H2O)! Bei einem Carbonatanteil 5,5 % (großteils Calciumcarbonat) und einem spez. Trockengewicht der Erde von 1000 kg/m3 auf 100 m2 bis in eine Tiefe von 0,5 m mit 0,055 · 50 0000 = 2750 kg Carbonatanteil muss man 2750 · 2,78 kg ≈ 7645 kg Eisen-(II)-sulfat in 100 m2 einarbeiten. |
Quantitativ überwiegen bei sauren Ackerböden wahrscheinlich die Ammoniumkationen (NH4+), bei alkalischen Böden die Nitratanionen (NO3− ). Können Pflanzen aufgrund der Durchlässigkeit der Wurzelmembranen nur NH4+ aufnehmen, dann sind sie auf saure Böden angewiesen. Wenn sie vorwiegend Nitrat (NO3−) aufnehmen, können sie nur auf basenreichen Böden wachsen. Die Ansprüche an den pH-Wert im Boden sind geringer, wenn die Wurzelmembranen sowohl Ammoniumionen als auch Nitrationen durchlassen. Wird mit Ammoniumnitrat (NH4NO3) gedüngt, so werden gleich viele Ammonium- und Nitrationen eingebracht. Der pH-Wert von Ammoniumnitrat wird in Abhängigkeit der Konzentration in Abschnitt 17.2.1 angegeben.
Ammoniumsulfat bindet das Calcium und scheidet es nicht mehr als Calcium aus, sondern es bleibt als Gips gebunden.
|
| CaCO3 : (NH4)2SO4 = 100,087 : 132,14 g/mol = 1 : 1,32 |
Ein kg Caliumkarbonat (Calzit - Kalkstein) kann mit 1,32 kg Ammoniumsulfat neutralisiert werden. Das resultierende Ammoniumcarbonat ist eine leichte Base.
| Stickstoffgehalt | pH–Wert | |
| Ammoniumsulfat | 21 % | stark senkend |
| Ammoniumnitrat | 35 % | schwach senkend |
| Kaliumnitrat | 15 % | neutral |
| Calciumnitrat | 17 % | anhebend |
| Schwach sauer wirkend sind Mehrnährstoffdünger mit höherem Ammonium- als Nitratanteil. Stark sauer wirkend sind Einzeldünger wie Ammoniumsulfat (schwefelsaurer Ammoniak). Alkalisch wirkend sind Mehrnährstoffdünger mit höherem Nitrat- als Ammoniumanteil, Einzeldünger wie Calciumnitrat (Kalksalpeter). |
Um in kalkhältigen Böden einen pH-Wert unter 7 zu erreichen, muss der Carbonatpuffer ausgeschaltet werden. Die folgende Tabelle mit Angaben über neutralisierend wirkende Zuschlagsstoffe wurde aus [R.G02] und [Mic03] entnommen. Ich möchte Anmerken, das diese Tabelle nur ganz grob die Verhältnisse aufzeigen kann, so kann je nach Nitrifizierung von Stickstoffdüngern die Neutralisierung stark schwanken. Organischen Substanzen variieren in ihrer aus Wirkung stark!
Die Werte zeigen gute Übereinstimmung mit den beiden Rechnungen aus Abschnitt 18.2.3.
| Bindungssubstanz | 1 kg CaCO3 wird durch |
| [kg] neutralisiert | |
| Schwefel (S) | 0,32 |
| Schwefel-ummantelter Harnstoff [38-0-0] | 0,85 |
| Ammoniumsulfat Dünger ((NH4)2SO4) [21-0-0] | 0,91 |
| Harnstoff ((NH2)2CO) [46-0-0] | 1,23 |
| Ammoniumnitrat Dünger (NH4NO3) [34-0-0] | 1,67 |
| 32% flüssiger Harnstoff-Ammoniumnitrat [32-0-0] | 1,82 |
| Diammoniumphosphat (18-46-0) | 1,43 |
| Aluminumsulfat (Al2(SO4)3) | 2,22 |
| Eisensulfat (Fe(SO4)3·9H2O) | 2,86 |
| Torf | 3,33+ |
| Pinienrinde | 5+ |
| Kompost | 10–∞ |
| anderer tierischer Dünger | 10–∞ |
| Geflügelmist | - |
| Andererseit versucht man mit der Kalkung von Äckern, Gärten und Wäldern, sowie strengeren Vorgaben zur Abluftreinigung, die Bodenversauerung einzudämmen. Mit der Aufbringung von kohlensaurem Kalk – Calciumkarbonat (CaCO3) – wird bis zu einem bestimmten Punkt die saure Wirkung aufgehoben. |
Kohlenstoffdioxid und Wasser bilden Kohlensäure (H2CO3), die dann wieder zu Hydrogencarbonat (HCO3−), einer schwachen Base und einem Wasserstoff-Proton (H+) dissoziiert:
| CO2 + H2O → H2CO3 → HCO3− + H+ |
Die Bildung von Kohlensäure (H2CO3) funktioniert bei einem pH-Wert zwischen 5 und 7 ??, da unterhalb von 5 keine Kohlensäure mehr gebildet wird.
So entstehen auf landwirtschaftlich genutzten Flächen etwa 10kg Wasserstoffionen (H+)/ha und Jahr – zu deren Neutralisation 500kg Calciumcarbonat (CaCO3)/ha benötigt werden [Wik15b].
Bei Eintrag von z.B. 100 kg einer Ammoniumverbindung (NH4+) in den Boden, können sich bis zu 7kg Wasserstoffionen/Protonen (H+)/ha durch Pflanzen bilden. Die kompensierende Kalkmenge beträgt 350 kg Calciumcarbonat (CaCO3) [Wik15b].
Die angegeben Tabelle soll einen Überblick geben, welche Baumarten auf welchen Böden gedeihen. Weiter kann man aus phänomenologischer Sicht, bei natürlich gewachsenen Baumbeständen, daraus ganz grob auf neue Pflanzmöglichkeiten schließen. Edelkastanien gedeihen in Mitteleuropa, wenn die klimatischen Bedingungen stimmen, auch meist auf Böden, die von Eichen genutzt werden. Diese vergleichenden Annahmen ersetzen natürlich keine Bodenanalyse.
| Art | lateinische Bez. | saure Böden | alkalische Böden | pH-Wert |
| Italienische Steinkiefer | Pinus pinea | X | X | 4,0-9,0 |
| Immergrüne Steineiche | Quercus ilex | X | X | 5,5-8,5 |
| Elsbeere | Sorbus torminalis | X | X | 6,0-8,5 |
| Österreichische Schwarzkiefer | Pinus nigra | X | X | 6,0-8,0 |
| Gemeine Esche | Fraxinus excelsior | X | X | 5,5-8,0 |
| Schwarznussbaum | Juglans nigra | X | X | 6,0-8,0 |
| Bergahorn | Acer pseudoplatanus | X | X | 6,0-8,0 |
| Zerr-Eiche | Quercus cerris | X | X | 5,0-8,0 |
| Rotbuche | Fagus sylvatica | X | 6,5-7,5 | |
| Rosskastanien | Aesculus hippocastanum | X | X | 6,0-7,5 |
| Sommerlinde | Tilia platyphyllos | X | X | 6,0-7,5 |
| Walnuss | Juglans regia | X | 6,5-7,5 | |
| Stieleiche | Quercus robur | X | 5,5- 7,0 | |
| Streu-Obstbäume | X | 6,0- 7,0 | ||
| Edelkastanie | Castanea sativa | X | 4,5- 6,5 | |
| Moorbirke | Betula pubescens | X | < 5,0 | |
Abrisse stooling, stool layering Ableger durch Anhäufeln der bis zum Wurzelstock zurückgeschnittenen Mutterpflanze ADP adenosine diphosphate Adenosindiphosphat, entsteht bei der Hydrolyse von ATP über ein Enzym und gibt Energie frei ATP adenosine triphosphate Adenosintriphosphat – ein Nukleotid, universeller Energieträger in Zellen Abmoosen air layering/marcotting ein Trieb wird zur Wurzelbildung mit Substrat – meist Moos – umwickelt, der Trieb ist nicht mit dem Erdboden verbunden. Adventivknospe adventitious bud ungewöhnlich dazugekommende Knospe am Stamm oder Wurzel Adventivwurzel adventitious root Wurzel, die nicht am Hauptvegetationpunkt entsteht, wie aus Kallusgewebe Altersdimorphismus age dimorphism Veränderung im Alter Amme über umgedr. Radicula inverted radicle method Amme wird über die umgedrehte Radikula verbunden Ammenvermehrung nurse seed grafting Vegetative Vermehrung mit Amme Ammoniak ammonia NH3 – stark stechend riechendes, farbloses, wasserlösliches und giftiges Gas Ammonium ammonium cation NH4+ positiv geladenes Ammoniumion Ammoniumnitrat ammonium nitrate NH4NO3 Anion anion negativ geladenes Ionen – wandert zur Anode Anemophilie anemophily Windblütigkeit – Windbestäubung Apikaldominanz apical dominance Unterdrückung des Wachstums von Seitentrieben. Gesteuert durch terminale Knospe über Auxin Auxin Auxine auxine Pflanzenhormon–Gruppe zur Zellstreckung, Wurzelbildung Axilläre Knospe axilla gemma axillary bud Seitenknospen (Achselknospen) am Trieb BAP 6-benzylaminopurine C12H11N5 – 6–Benzylaminopurin ist ein Zytokin Buchengewächse Fagaceae beech Calciumcarbonat calcium carbonate CaCO3 – kohlensaurer Kalk, Calzit Calciumhydroxid slaked lime Ca(OH)2 – gelöschter Kalk Calciumoxid burned lime/quicklime CaO – ungelöschter Kalk, Ätzkalk Calciumsulfat calcium sulfate CaSO4 – wasserunlöslich Chelat chelate Chelatkomplexe – machen Düngerelemente wie Eisen leicht löslich Chlorit chlorite Vierschichtsilikat Chymosin chymosin Lab-Enzym Diammoniumphosphat diammonium phosp. (NH4)2HPO4 – (DAP) Diammoniumhydrogenphosphat ist ein Ammoniumsalz Doppelsuperphosphat dicalcium phosphate Ca(H2PO4)2 (35 % P2O5) Dormanz dormancy alle Formen Entwicklungsverzögerung, z.B. Keimruhe (Winterruhe) Edelauge bud Knospe einer wertvollen Sorte Edelkastanie amer. Castanea dentata sweet chestnut amerikanische Edelkastanie Edelkastanie chin. Castanea mollissima chinese chestnut chinesische Edelkastanie Edelkastanie eur. Castanea sativa eur. chestnut europäische Edelkastanie Edelkastanie jap. Castanea crenata jap. chestnut japanische Edelkastanie Edelreis scion abgeschnittene einjährige Triebe einer wertvollen Sorte Einschnürring girdle, metal ring Ring am Trieb um den Phloemstrom weitgehend zu unterbinden um die Bewurzelung oberhalb der Einschnürrstelle anzuregen Endständige Knospe terminal bud Knospe am Ende des Triebes Entomophilie entomophily Pollen werden durch Insekten verteilt Epykotyl Epycotyl epicotyl Abschnitt von Kotyledonen zum ersten Folgeblatt Fruchtbecher Cupula cupula Fruchtbecher Glucose glucose C6H12O6 – Traubenzucker Harnstoff Urea urea CH4N2O – Endprodukt eines Stoffwechsel, weißer, kristalliner Feststoff Hochmoortorf Spagnum pet moos abgestorbenes Sphagnum Moos Homozygotie homozygous Reinerbigkeit Hydrogencarbonat hydrogen carbonate HCO3− – Hydrogencarbonatanion (alt Bicarbonat), Salze der Kohlensäure Hydrolyse hydrolysis ist die Spaltung einer (bio)chemischen Verbindung durch Reaktion mit Wasser Hydroxid hyroxide OH− – Hydroxidanion; entstehen, wenn Basen mit Wasser reagieren Hypokotyl Hypocotyl hypocotyl Abschnitt von Radikula zu den Kotyledonen IAA indole-3-acetic acid C10H9NO2 – Indol-3-essigsäure auch nat. vorkommendes Auxin IBA indole-3-butyric acid C12H13NO2 – 4-(Indol-3-yl)buttersäure Auxin Illit illite Dreischichtsilikat KAK cation–exchange capacity Kationenaustauschkapazität des Bodens Kallus Callus callus Wundgewebe der Pflanze, undifferenzierte Zellkomplexe Kambium Cambium cambium Wachstumsschicht zw. der Splintholzzone und Rinde Kaolinit kaolinite Porzellanerde – Zweischichtsilikat Kation cation positiv geladenes Ionen – wandert zur Kathode Knoten/Nodium Nodus node Verdickter Blattansatzpunkt an Ast/Sprossachse Kohlensäure carbonic acid H2CO3, Reaktionsprodukt von Kohlenstoffdioxid mit Wasser. Kotyledon Kotyledon cotyledon Keimblatt Lehm Lutum loam Gemisch aus etwa 40 % Ton + 40 % feinem Sand + 20 % Schluff (Korngr. zw Sand und Ton) Leitgewebe vascular bundle im Xylem + Phloem Löss loess besteht hauptsächlich aus Schluff – ist sandiger als Lehm und 8–20% Kalk Meristem Meristem meristem Bildungsgewebe aus undifferenzierten Zellen der Pflanzen – Stammzellen Montmorillonit montmorillonite häufiges Dreischichtsilikat Monözie monoclinous Einhäusigkeit, männl. und weibl. Büten auf der gleichen Pflanze NAA 1-naphthaleneacetic acid C12H10O2 – 1 Naphthylessigsäure – Auxin Nitrat nitrate NO3− – Nitratanion, Salze der Salpetersäure (HNO3): NaNO3, KNO3, NH4NO3 Nitrit nitrite NO2− – Nitritanion, Salze der salpetrigen Säure (HNO2), Pökelsalz: Kaliumnitrit, Natriumnitrit, Natriumnitrat Nukleotid nukleotide ist ein Grundbaustein von Nukleinsäuren, also Desoxyribonukleinsäure (DNA) und Ribonukleinsäure (RNA) Perikarp Pericarp pericarp Gehäuse Phloem phloem Teil des Leitgewebes für Transport von Wasser, Saccharosen und Aminosäuren von den Blättern in basaler Richtung Pilz Cryphonectria parasitica chestnut blight Schlauchpilz, bewirkt Kastanienrindenkrebs Pilz Phloeospora castanicola bewirkt Sprühfleckenkrankheit Pilz Phytophthora cinnamomi dieback bewirkt Tintenkrankheit Pilz–Hyphen fadenförmige Zellen der Pilze Pilz–Mycel ist die Gesamtheit aller Hyphen, normalerw. unter der Erde Radikula Radicula radicle Keim– oder Pfahlwurzel Rosskastanie Aesculus hippocastanum [horse] chestnut Rotbuche Fagus sylvatica beech Salmiak ammonium chloride NH4Cl – Ammoniumchlorid, farbloser, kristalliner Feststoff Schössling frischer Austrieb einer Pflanze Smectit smectite Dreischichtsilikat Sprossachse plant stem Verbindung zw. Wurzel und Blätter – Stamm mit Ästen Stamen stamen das Staubblatt, Staubgefäß ist das pollenerzeugende Organ der Blüte Steckling cuttings Abgeschnittene Triebe, die zum Bewurzeln in ein Substrat gesetzt werden Steineiche Quercus ilex evergreen oak Superphosphat monocalcium phosphate besteht aus Ca(H2PO4)2 und wasserunlösslichem CaSO4· 2 H2O (16 – 22 % nP2O5) Symbiose–Pilz Ekto-Mykorrhiza Pilz symbiotic fungus geeignete Symbiose–Pilzgruppe für Laubbäume sind Ekta–Mykorrhiza Sympodium sympodium Endknospen, die im Herbst absterben Ton Argilla clay enthält Schichtsilikate wie Illit, Montmorillonit oder Kaolinit Unterlage rootstock besteht aus Wurzeln und Stamm, wird beim Veredeln verwendet Veredelung Copulare grafting Vegetative Vermehrungsart (Kopulation/Okulation) Vermiculit vermiculite Dreischichtsilikat Wasserschoss (epicormic shoot) Wassertrieb – Sommertrieb ausgehend von altem Holz aus einem schlafendem Auge – ein aufrechter Trieb mit relativ weichem Gewebe Xylem Xylem xylem Teil des Leitgewebes im Kambium für den Transport von Wasser und anorg. Nährsalzen zu den Blättern Zygote Zygote zygote/proembryo befruchtete Eizelle Zytokin Cytokine cytokine Pflanzenhormon–Gruppe zum Wachstum und Differenzierung verderbliche Samen recalcitrant/perishable se. Samen mit schlechter Lagerfähigkeit an der Spitze liegend apikal apical z.B. Spitze des Triebes betreffend die Basis bildend basal basal z.B. unteres Triebende am Ende gelegen terminal terminal z.B. die Triebspitze betreffend
Für Fachbegriffe der botanischen Literatur ist [Wik15a] eine gute Basis.
This document was translated from LATEX by HEVEA.